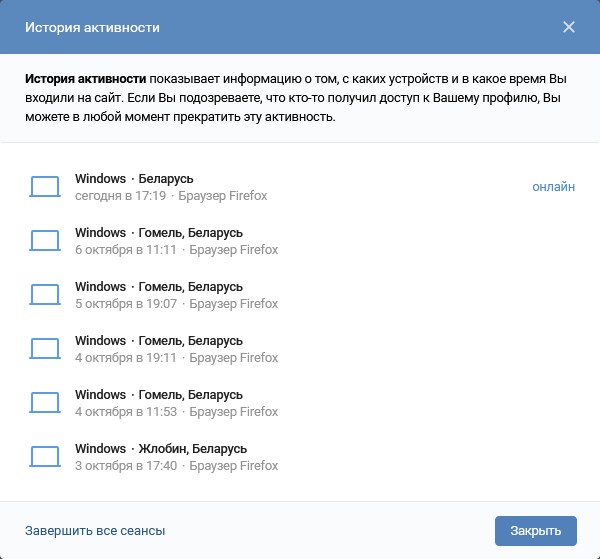
Почему сайт ВКонтакте пишет теперь вместо «1000» просмотров «1К»? — Обсуждай
Почему сайт ВКонтакте пишет теперь вместо «1000» просмотров «1К»? — ОбсуждайFB
Foxy Beige
Почему сайт ВКонтакте пишет теперь вместо «1000» просмотров «1К»? сайт вконтакте просмотр
1029
83
4
Ответы
Се
Сергей
Почему k это тысяча нужно искать в Древней Греции, а точнее — в системе древнегреческого языка. В нём существовало уникальное слово «Chilioi» — оно использовалось для обозначения тысячи, греческое Chilioi перешло во французский язык, далее — французы сократили его до простого «кило». В старославянской азбуке К-20, эти южане привыкли всё преувеличивать, вот и результат. .
.
0
Ва
Валерий
Это 10 в кубе. Это обозначение давно используется на просторах инета. А пошло скорее всего с начала компьютеризации. Так проще и короче записывать числа. Объёмы памяти были не большие, от туда пошло.
0
ЮШ
Юрий Шокин
Буква «К» означает кратность (К) = 1000 ед. измерения. Например : килоВОЛЬТ= 1000 вольт, килоГРАММ=1000 грамм, килоМЕТР=1000 метрам, килоОМ=1000 ом, килоАМПЕР=1000 ампер…..и так далее.
0
ВС
Виталий Соколов
мода пошла из рэп-тусовки, где проще говорить фразы типа «два КА один восемь» вместо «две тысячи восемьнадцатого года».
8 · Хороший ответ
13
0
Ильмир Шарифуллин
Для страждущих ответа, вот вам Авторитетный Источник™
http://ru.wikipedia.org/wiki/%D0%9F%…8_%D0%A1%D0%98-очень подробный ответ
0
ВА
Виктор Александров
потому что 1к это 1 килобайт. Килобайт — единица измерения объема передаваемой или хранимой информации
0
СК
Сергей Короленко
0
VN
Vita Nagib
К -кило -это тысяча или 10 в 3-ей степени ; М -мега миллион 10 в 6-йстепени
0
ТП
Толя Попов
а. …………..хх….его знаетсам об этом думаю………день и ночь
…………..хх….его знаетсам об этом думаю………день и ночь
0
ВМ
Владимир Малиновский
Вот не знаю… Из экономии, возможно. В «1К» знаков меньше.
0
ВЦ
Виктор Царук
В системе мер Кило-тысяча.Мега-Миллион.Тера-……и.т.д.
0
Alexander Zlobin
Почта на яндпксе тоже так пишет и Одноклассники.
0
Кс
Ксения™
Ахаха не думаю что здесь поймут… Не в теме)
Не в теме)
0
FB
Foxy Beige
Почему?)
1
ВЛ
Виктор Луговской
Они сейчас в Кельвинах мерят накал страстей.
0
СК
Сергей Константинов
Все так стали писать.Нули на клаве клинят
0
H_
Hsd_T _
1к = 1000
Мы находимся в эпоху аббревиатур
0
Сергей Радиевич Фасулати
Наверно, это означает «1 килопросмотр»
0
FB
Foxy BeigeХа-ха. Ха-ха-ха.
Ха-ха-ха.
1
Та
Татьяна
Дурная мода. Своего же ума не хватает.
0
Дима Жидак
так сайт решил и их основатели сайта
0
ВМ
Владимир Морозов
Думаю что К — это кило, то есть тысяча
0
Следующая страница
Охват Вконтакте: виральный, подписчиков, полный, рекламный
Реклама в соцсетях – это эффективный способ достучаться до целевой аудитории. Поэтому многие бренды развивают свои сообщества в ВК, вкладывая в раскрутку время и деньги.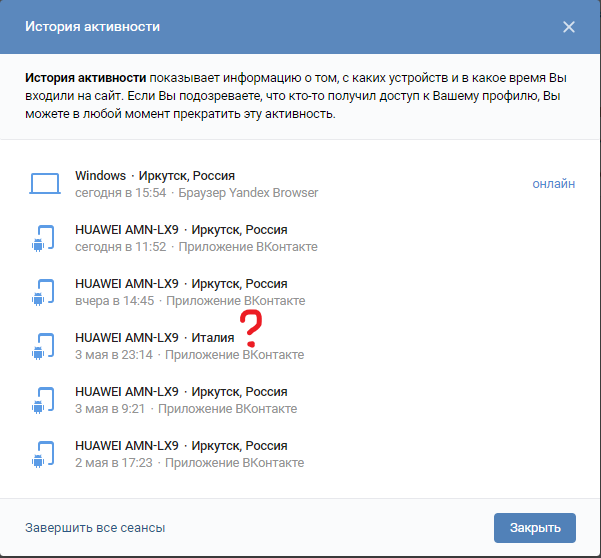
Не всегда получается добиться высокой активности аудитории. Проблема может быть в том, что подписчики не видят постов сообщества в своей ленте. Чтобы исправить положение, необходимо позаботиться об увеличении вирального охвата подписчиков Вконтакте.
Содержание
- 1 Полный охват
- 2 Виральный охват
- 3 Охват подписчиков
- 4 Рекламный охват
- 5 Как посмотреть охват
- 6 Как повысить охват
- 6.1 Создаём контент-план
- 6.2 Цепляющие посты
- 6.3 Лайки
- 6.4 Задаём вопросы
- 6.5 Получаем комментарии
- 6.6 Публикуем видео
Полный охват
Это все виды охвата. Состоит из:
- вирального;
- охвата подписчиков;
- рекламного.
То есть, это максимальное количество просмотров поста, как уникальных, так и не уникальных. Даже если человек промотал в своей новостной ленте запись, не просматривая, это принесёт +1 к полному охвату группы.
Виральный охват
Называют количество пользователей, которые не подписаны на сообщество, но увидели пост на стене или в новостях. Этот показатель измеряет количество уникальных пользователей и общих просмотров записи.
Этот показатель измеряет количество уникальных пользователей и общих просмотров записи.
Например, пользователь сделал репост со стены друга. Просмотр контента подписчиками и друзьями человека будет виральным по отношению к первоисточнику.
Если говорить о коммерческих постах, то виральным будет охват той аудитории, которая не указывалась в настройках объявления. Рекламные записи создаются для демонстрации пользователям, не подписанным на паблик ВК.
Если представитель целевой аудитории бренда делает репост этой записи, то её увидят люди, которые не соответствуют настройкам показа рекламного поста. Просмотры рекламодатель получает бесплатно.
«Ушло в народ» – высказывание относится к виральному контенту. Значит, информация хорошо принята и распространена аудиторией. Создаётся сетевой эффект, контент становится вирусным и дешёвым.
Охват подписчиков
Это количество пользователей, которые просмотрели запись группы ВК. Они разделяются на две категории:
- Подписчики – увидели новый пост в ленте.

- Не подписчики – увидели запись на странице сообщества в ВК.

При этом охват подписчиков и количество просмотров – не одно и то же. Многие владельцы групп в ВК путают эти понятия.
Просмотры свидетельствуют о том, сколько раз страница или запись была просмотрена. Система учитывает все просмотры. Пользователь может увидеть пост в сообществе и новостной ленте несколько раз, и каждый из них пойдёт в зачёт.
Рекламный охват
Показывает количество людей, которые просмотрели контент, благодаря рекламным настройкам. Это следствие работы таргетированной рекламы, которая работает с целевой аудиторией по заданным параметрам. В ходе поиска учитываются интересы пользователей, их возраст, пол.
Как посмотреть охват
Владельцы пабликов в ВК хотят знать охват своих сообществ. Эта информация не засекречена, и получить к ней доступ можно всего в несколько кликов мышки.
Чтобы узнать данные для сообществ:
- Найти кнопку «Статистика» на панели управления.
- Переходим в раздел «Охват», где можно посмотреть все данные о популярности постов.

Владельцам бизнес-профилей и блогерам будет интересно узнать статистику аккаунта. Нажмите кнопку статистики, расположенную сразу под аватаркой.
Эти данные доступны всем пользователям, имеющим более 50 подписчиков. Если же подписчиков больше сотни, то можно получить доступ к более подробному анализу, где, кроме охвата, будет возраст, пол и страны посетителей страницы.
Как повысить охват
У владельцев групп возникает следующая ситуация: большая аудитория и регулярный выход постов, но нет активности. В ВК ввели умную ленту, которая фильтрует контент. Теперь посты могут видеть не все подписчики сообщества.
Система выполняет тщательный анализ активности пользователя. Показывается контент, который может вызвать интерес. Поэтому над повышением охвата поста без накрутки придётся поработать.
Создаём контент-план
Чтобы посты повышали активность в группе ВК, их выкладывают по тщательно проработанным алгоритмам. Выход публикаций привязывается ко времени, дате, теме в установленной последовательности.
Выход публикаций привязывается ко времени, дате, теме в установленной последовательности.
Создавая контент-план, проанализируйте целевую аудиторию. Изучите интересы пользователей, их восприятие информации.
Скачайте шаблон контент-плана, отредактируйте его или создайте новый.
Цепляющие посты
Подписчик видит публикацию двумя способами: в сообществе или новостной ленте. В обоих случаях пост должен привлечь внимание пользователя ВК. Если же этого не произойдёт, то не будет лайков, репостов и комментариев. Основная задача контента – выделиться в ленте, «зацепить». Найдите то, что интересует вашу аудиторию.
Лайки
Стимулируйте подписчиков ставить лайки. Для этого размещайте интересные посты. Регулярно выкладывая публикации, вы прививаете пользователям привычку лайкать и репостить.
Заслуживают одобрения посты с кейсами. Ставить лайк, чтобы сохранить в закладках публикацию – это распространённая среди пользователей привычка. Публикации не должны отличаться от тематики сообщества.
Часто подписчики не ставят первый лайк. После публикации лайкните её сами, попросите друзей сделать то же самое. Такие действия поднимут контент в глазах целевой аудитории без накруток.
Задаём вопросы
Для повышения активности подписчиков Вконтакте используйте опросы. Эта схема работает потому, что людям интересно не столько ответить на вопрос, сколько узнать, что ответили другие. Придумайте красивое оформление, завлекающую тему и опубликуйте.
Получаем комментарии
Подписчики редко комментируют публикации по своей инициативе. Рекомендуем задавать вопросы по теме поста. Подписчики с удовольствием делятся личным опытом или мнением в комментариях.
По аналогии с лайками оставляйте первый комментарий. Напишите его реалистично.
Для активного обсуждения создаётся публикация на провокационную тему. Однако не стоит злоупотреблять таким контентом, он может отвлечь внимание пользователей от тематики группы.
Публикуем видео
Наибольший охват получает видеоконтент. Чтобы умная лента Вконтакте «рассказала» о публикации подписчикам, заливаем видео в раздел «Видеозаписи», а после – выкладываем отдельным постом. Если же запись взята из стороннего ресурса, то лента уменьшает число показов.
Чтобы умная лента Вконтакте «рассказала» о публикации подписчикам, заливаем видео в раздел «Видеозаписи», а после – выкладываем отдельным постом. Если же запись взята из стороннего ресурса, то лента уменьшает число показов.
Раскрутка сообщества в ВК – это трудоёмкий процесс, но при верном подходе получится вывести сообщество в топ. Пользуйтесь советами и стимулируйте активность подписчиков. Группа с активной аудиторией привлекает большее число пользователей.
что дальше? — Маркетинг на vc.ru
Специалисты digital-агентства Nimax и сервиса аналитики соцсетей LiveDune рассказали, на какие метрики обратить внимание при продвижении на новой площадке.
2339 просмотров
Долгое время основной площадкой для компаний и брендов был Instagram*. В пуле возможностей у Вконтакте есть все опции Instagram*, а также большой потенциал и огромное количество инструментов под различные потребности. В этой статье сосредоточимся на метриках и аналитике.
Следим за метриками
Несмотря на широкий инструментарий, статистика сообщества, которую предоставляет Вконтакте скромная и не самая удобная: цифры есть, но делать выводы по ним сложно. Поэтому в Nimax мы анализируем статистику сообществ в LiveDune. Сервис показывает не просто таблицы и графики, он помогает принимать решения о контенте и продвижении с опорой на цифры: о чем писать посты, когда их публиковать, как эффективно продвигать сообщество и получать отдачу.
Поэтому в Nimax мы анализируем статистику сообществ в LiveDune. Сервис показывает не просто таблицы и графики, он помогает принимать решения о контенте и продвижении с опорой на цифры: о чем писать посты, когда их публиковать, как эффективно продвигать сообщество и получать отдачу.
Рассмотрим метрики, на которые стоит обратить внимание при анализе сообщества Вконтакте.
Охват
Компании, бренды, специалисты по маркетингу и контент-менеджеры тщательно следят за охватом. Эта метрика показывает количество уникальных пользователей, которые увидели публикацию. Вконтакте не считает охват для рекламных объявлений совокупно за весь период показов или на уровне кампании. А для сообщества и публикаций в ленте делает это не очень удобно. Это одна из преград в работе для тех, кто привык опираться на показатель охвата в Instagram*.
В ВК охват можно посмотреть во внутренней статистике сообщества в одноименном разделе. График не содержит выводов и неудобен для анализа. Например, если нужны суммарный охват за определенный период, придется делать выгрузку в excel и дополнительно складывать все данные по дням. Слишком много действий, чтобы посмотреть всего один показатель. А также это не совсем корректный подход: суммируя охват, мы не можем исключить попадания в выборку одних и тех же пользователей.
Например, если нужны суммарный охват за определенный период, придется делать выгрузку в excel и дополнительно складывать все данные по дням. Слишком много действий, чтобы посмотреть всего один показатель. А также это не совсем корректный подход: суммируя охват, мы не можем исключить попадания в выборку одних и тех же пользователей.
LiveDune экономит время и наглядно отображает данные, что позволяет быстро принимать решения.
- Все графики содержат краткие выводы. Вы сразу сможете посмотреть суммарный и средний (среднее арифметическое и по медиане) охват за месяц.
- Сервис не только показывает данные за выбранный вами временной отрезок но и всегда сравнивает их с аналогичным периодом в прошлом. Например, вы смотрите охват с 10 по 20 марта и сразу можете узнать, как изменился этот показатель в сравнении с периодом с 10 по 20 февраля.
- Если заметили резкий рост или спад на графике, то можете посмотреть, сколько и какие посты выходили в дни пиковых значений. Просто нажмите на нужный день на графике — появится список опубликованных постов. Это поможет вам понять, какой контент повлиял на изменение значений.
 Просто нажмите на нужный день на графике — появится список опубликованных постов. Это поможет вам понять, какой контент повлиял на изменение значений.
Просто нажмите на нужный день на графике — появится список опубликованных постов. Это поможет вам понять, какой контент повлиял на изменение значений.Вовлеченность
Следующая по важности метрика — вовлеченность. Она показывает не просто объем уникальных пользователей, которые видели ваши посты, но степень их погружения в эти публикации: нравится ли им читать ваши материалы, хочется ли их обсудить или поделиться ими с друзьями. Колебания метрики помогают понять настроение и интересы аудитории.
Вовлеченность пользователей LiveDune позволяет анализировать по нескольким метрикам: ER, ERR, ERV, RR.
Для чего так много показателей? Полнота данных помогает сделать верные выводы. Разберемся на примере. Среднее количество реакций относительно подписчиков (ER) — привычная метрика для SMM-менеджера или таргетолога. В динамике вы можете увидеть, как этот показатель падает, и огорчиться. И даже решить, что тематика постов подобрана не верно.
Но снижение показателя может быть связано с ростом подписчиков сообщества. То же количество реакций вы теперь делите на большее число подписчиков. Как так получилось? Ваша аудитория выросла, но не видит в ленте публикаций. Поэтому не оставляет лайки, не пишет комментарии и не делится постами с друзьями. Это подтвердит следующий показатель — количество реакций относительно охвата постов (ERR).
Здесь количество реакций делим не на всех пользователей, а на тех, кто видел ваши публикации. И этот показатель вырос. Значит, аудитории нравятся посты, которые они видят в вашем сообществе. Получается, коммуникационная стратегия подобрана верно. Здесь на помощь SMM-менеджеру приходит таргетолог, который покажет лучшие публикации большему числу пользователей, и увеличит охват постов.
Наш подход — не делать поспешных выводов, не опираться на ощущения, а полагаться на цифры
Маша Курагодникова
Анализ контента
Еще одно слабое место внутренней статистики сообщества Вконтакте — анализ контента. Во-первых, данные будет доступны для сообществ только с количеством подписчиков от 5000. Во-вторых, для анализа контента будут доступны всего три метрики: лайки, репосты, комментарии.
Во-первых, данные будет доступны для сообществ только с количеством подписчиков от 5000. Во-вторых, для анализа контента будут доступны всего три метрики: лайки, репосты, комментарии.
Контент тоже анализируем в LiveDune: сервис собирает статистику даже для сообществ с менее чем 5000 подписчиков. Для анализа предоставляет данные по 11 метрикам. Таблицу можно сортировать по любому значению: по убыванию или возрастанию.
Также у LiveDune есть собственная метрика для анализа контента — оценка постов. Система оценивает реакцию подписчиков на публикации, опираясь на скорость набора лайков и исторические данные — как пользователи реагировали на предыдущие посты. Если пользователям нравится публикации, они активно взаимодействуют с ней, тем выше будет оценка. Если же пользователи неохотно ставят лайки по сравнению с предыдущими постами, то оценка будет ниже. Посты с самой высокой оценкой «А+» можно смело ставить на продвижение.
Самый популярный и простой способ анализировать контент — выстраивать ТОПы и анти-ТОПы по разным метрикам.
Аня Чернова
 Сравнивать худшие посты с лучшими и пытаться понять, как им можно доработать. Такой анализ в LiveDune можно делать по суммарному количеству реакций на посте, лайкам, комментариям, репостам, голосам, просмотрам, охватам,переходам, RR, ER, ERR
Сравнивать худшие посты с лучшими и пытаться понять, как им можно доработать. Такой анализ в LiveDune можно делать по суммарному количеству реакций на посте, лайкам, комментариям, репостам, голосам, просмотрам, охватам,переходам, RR, ER, ERRЕще один полезный инструмент — оценка публикаций в разрезе рубрик. Такой анализ — хорошее подспорьем при составлении контент-плана. Вы увидите, какие темы больше нравятся пользователям, а какие не находят отклика у аудитории. Также можно тестировать новые темы и оценивать их эффективность относительно устоявшихся.
Анализ контента по рубрикам помогает найти темы, на которые нет смысла тратить время и ресурсы. Часто бывает так, что какой-то отдельно взятый пост из рубрики выстрелил, и мы принимаем решение создавать такой контент дальше. Но если посмотреть на эффективность рубрики в целом, картина может быть совершенно противоположной. Или со временем рубрика просто приестся аудитории.
Аня Чернова
Еще один пункт в копилку оптимизации работы — возможность выстраивать ТОПы и анти-ТОПы постов по конкретным метрикам. Быстрый анализ в сводной таблице выглядит так:
Анализ конкурентов
Сравнительную таблицу по нескольким сообществам конкурентов в LiveDune можно собрать за несколько кликов. Она поможет вам сориентироваться на местности:
LiveDune — сервис не только для аналитики соцсетей, но и автоматизации SMM-процессов. Например, здесь можно обрабатывать комментарии и сообщения, планировать публикации постов, отслеживать выполнение KPI.
В Nimax мы любим LiveDune еще и за то, что все данные удобно выгружаются в отчет. Для своих клиентов мы готовим промежуточные сводки еженедельно и делаем общие выводы каждый месяц. Команда не тратит время на сбор статистики, а сразу приступает к компиляции данных и их анализу, проверяет поставленные гипотезы, корректирует контент-план и план по продвижению.
Полезными материалами о маркетинге в новом времени мы делимся в телеграм-канале Команда Nimax. Будем рады увидеться в комментариях. Остаемся на связи!
Будем рады увидеться в комментариях. Остаемся на связи!
* Facebook и Instagram запрещены на территории России. Компания-владелец Meta признана экстремистской организацией.
Как сделать карточку музыканта в ВК, Spotify, Apple Music?
Популярность стриминговых сервисов за 2020 год приросла среди музыкантов и слушателей. Шума добавил выход Spotify в России. Да и людей, которые активно занялись музыкой добавилось.
Первое, что видит человек, если он заинтересован каким-то артистом — это карточка музыканта. В некоторых стриминговых сервисах карточку называют профилем.
Сегодня разберем тему о том, как создать карточку музыканта на трех основных стриминговых площадках России: VK/BOOM, Spotify, Apple Music.
Что такое карточка музыканта?
Это страница, на которой собраны все песни и альбомы музыканта.
Здесь можно найти ссылки на официальную группу или паблик, или личный профиль артиста в соцсетях. Здесь же можно разместить фотографии и биографию исполнителя.
Создание карточки музыканта чаще всего сопряжено с первой заливкой треков и регистрацией в сервисе. Расскажем об этом по порядку.
Создание карточки в ВК/BOOM
В приложении BOOM карточка музыканта формируется с сайта VK, так как BOOM — это просто аудиозаписи из Вконтакте, вынесенные в отдельное приложение.
Карточка создается автоматически при загрузке первых треков на площадку VK/BOOM.
Создать вручную карточку музыканта нельзя.
Если вы уже загружали свою музыку через лейбл или дистрибьютора, то у вас уже должна быть карточка музыканта. Просто введите имя исполнителя в поиске по аудиозаписи.
Как привязать к паблику и разместить свою фотографию?Для этого понадобится два фото: для десктопной и мобильной версий.
- Десктопная версия: размером 1510*344 px.
- Мобильная версия: размером 1440*730 px.
Чтобы добавить фотографии к карточке достаточно написать в службу технической поддержки. Если фото соответствует требованиям, его разместят.
Если фото соответствует требованиям, его разместят.
Чтобы привязать карточку музыканта к паблику, необходимо так же писать в техподдержку. Для этого название паблика должно совпадать с названием музыканта. Еще несколько условий: создателем паблика должен являться сам исполнитель, а все администраторы должны включить «Подтверждение входа» (2FA) в настройках безопасности страницы — с помощью аккаунты администраторов и самое сообщество получает дополнительный уровень защиты.
Если вы работаете с агрегатором или лейблом и по какой-то причине карточки музыканта по-прежнему нет, хотя вы уже заливаете треки во вКонтакте, пишите в техподдержку. В запросе обязательно указывайте название дистрибьютора, сценический псевдоним, названия релизов и уникальные UPS или ISRC-коды релизов.
Создание карточки в Spotify
В Spotify карточка артиста создается так же, как и в VK — автоматически. НО! Для этого необходимо зарегистрироваться в Spotify for Artists. И вот этот вопрос вызывает наибольшее количество вопросов.
Ниже приводим алгоритм регистрации в Spotify for Artists, если вдруг вы еще там не зарегистрированы:
- Регистрируйтесь в Spotify как слушатель и создайте аккаунт:
2. Переходите по ссылке: https://artists.spotify.com/
3. Нажимайте на Get Access в правом верхнем углу.
4. На странице будет выбор из двух вариантов. Сделайте этот выбор:
- Artist or manager — Артист или Менеджер
- Label team member — Член команды лейбла
5. На следующей странице Spotify предложит вам заявить права на профиль исполнителя. Смело жмите «Продолжить», чтобы получить доступ к Spotify для артистов из России.
6. В окне «What profile are you claiming? (На какой профиль вы претендуете?)» введите имя исполнителя или URL-адрес плейлиста.
7. Заполните информацию о себе «Tell us about yourself»
8. Ожидайте одобрение запроса от Spotify. В среднем срок ожидания занимает не больше суток.
Личный кабинет формируется автоматически и вся информация об артисте отображается во вкладке профиль (Profile): меняйте обложки, добавляйте биографию. Вы можете менять поле Artist’s Pick: размещать в нем трек, плейлист, альбом, гастроли или подкаст с добавлением небольшого сообщения для фанатов — чаще всего это поле используется для анонсов, но можно выдумать и какой-нибудь особый способ коммуникации через с аудиторией через это небольшое окошко.
В разделе ABOUT обязательно создайте фото галерею и добавлять до 100 фотографий. Поле для написания биографии артиста и ссылки на его соцсети так же размещаются здесь.
Создание карточки в Apple MusicВ Apple Music есть схожий со Spotify персональный кабинет или, как мы его называем, карточка музыканта. И ее не нужно каким-то особым способом создавать. В Apple Music нужно сделать одно действие: зарегистрировать Apple ID после чего просто получить доступ, что вызывает наибольшие трудности. Особенно, если нет Apple ID.
Особенно, если нет Apple ID.
Если у вас действительно нет Apple ID, то здесь, на официальном сайте яблочной техподдержки, есть четкая пошаговая инструкция для любого устройства. Самый простой способ создать Apple ID — «Создание идентификатора Apple ID на веб-сайте». Просто следуйте инструкции.
Чтобы зарегистрировать страницу артиста, войдите с помощью Apple ID в Apple Music for Artists или скачайте приложение.
Ссылка для входа в Apple Music: Apple Music for Artists.
1. Найдите «Запросить доступ к артисту» и укажите свою ссылку на страницу артиста в iTunes Store. Для этого в iTunes Store просто введите в поисковике имя артиста или название группы, выберите одни из альбомов или треков. Перепроверьте, что вы регистрируете нужного артиста.
2. Выберите роль и заполните необходимые поля в заявке. Чтобы ускорить одобрение своей заявки, войдите в свои профили соцсетей или в учетную запись своего дистрибьютора. Чем больше информации вы сможете предоставить. тем быстрее Apple Music for Artists подтвердит вашу личность и принадлежность к артисту.
тем быстрее Apple Music for Artists подтвердит вашу личность и принадлежность к артисту.
3. Проверка статуса заявки
Статус можно посмотреть в самом профиле, где будет указано: «На рассмотрении». При изменении статуса сервис сам сообщит об этом по электронной почте.
Иногда требуется дать доступ к странице артиста нескольким лицам: участникам группы, менеджеру артиста, представителям лейбла. Соответствующие функции есть в профиле.
Тысячи начинающих музыкантов зарегистрированы на разных сервисах и классное оформление карточки музыканта — это +1 к вашему успеху, ведь это важная составляющая, которая может много рассказать о вас, как о человеке.
Именно поэтому важно делать качественные и цепляющие обложки к своим трекам, искать возможности для крутых фотосессий, чтобы в любом профиле фанат, представители СМИ или лейбла видели вас, а не боролись с раздражением от низкого разрешения ваших фото. То же касается и текста вашей биографии — он должен быть информативным, цепляющим, ярким.
Apple Music spotify продвижение исполнителя продвижение музыки Стриминг
Квалификационные требования к спортивным судьям по виду спорта
Утверждены
приказом Министерства спорта
Российской Федерации
от 9 ноября 2020 г. N 822
КВАЛИФИКАЦИОННЫЕ ТРЕБОВАНИЯ
К СПОРТИВНЫМ СУДЬЯМ ПО ВИДУ СПОРТА «КОННЫЙ СПОРТ»
1. Требования для присвоения, подтверждения
квалификационных категорий спортивных судей
1.1. Требования к прохождению практики судейства
для присвоения квалификационных категорий спортивных судей
Наименование квалификационной категории спортивного судьи | Период прохождения практики судейства | Наименования должностей спортивных судей, предусмотренных правилами вида спорта | Статус и наименование соревнований, их количество | ||||||||||||||
Чемпионат России | Кубок России | Первенство России, Всероссийская Спартакиада между субъектами Российской Федерации | Другие всероссийские спортивные соревнования, включенные в ЕКП | Чемпионат федерального округа, двух и более федеральных округов, чемпионаты г. | Первенство федерального округа, двух и более федеральных округов, первенства г. Москвы, г. Санкт-Петербурга | Другие межрегиональные соревнования, включенные в ЕКП | Чемпионат субъекта Российской Федерации (кроме г. Москвы и г. Санкт-Петербурга) | Кубок субъекта Российской Федерации | Первенство субъекта Российской Федерации (кроме г. Москвы и г. Санкт-Петербурга) | Другие официальные спортивные соревнования субъекта Российской Федерации | Чемпионат муниципального образования | Первенство муниципального образования | Другие официальные спортивные соревнования муниципального образования | ||||
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | |
ВК | 3 года | ГСК | Главный судья | 4 | |||||||||||||
Заместитель главного судьи | |||||||||||||||||
Главный секретарь | |||||||||||||||||
Заместитель главного секретаря | |||||||||||||||||
Курс-дизайнер | |||||||||||||||||
Шеф-стюард | |||||||||||||||||
Заместитель шеф-стюарда | |||||||||||||||||
Технический делегат | |||||||||||||||||
Судья-член Гранд-Жюри | 2 | ||||||||||||||||
Ассистент технического делегата | |||||||||||||||||
Ассистент курс-дизайнера | |||||||||||||||||
Ассистент шеф-стюарда | |||||||||||||||||
Ассистент главного секретаря | |||||||||||||||||
Требования, обусловленные особенностями судейства | Для присвоения ВК необходимо в течение 3-х лет выполнить требования к прохождению практики судейства не менее 6-ти раз на соревнованиях соответствующего статуса, из которых не менее 2-х всероссийских соревнований, в указанных должностях спортивного судьи | ||||||||||||||||
Требования для кандидатов, имеющих РК или ВС | Для присвоения ВК необходимо выполнить требования к прохождению теоретической подготовки (участие во всероссийском семинаре) и требования к сдаче квалификационного зачета (сдать квалификационный зачет) без выполнения требований к практике судейства | ||||||||||||||||
1К | 2 года | ГСК | Главный секретарь | 5 | |||||||||||||
Заместитель главного секретаря | |||||||||||||||||
Судья-член Гранд-Жюри | |||||||||||||||||
Курс-дизайнер | |||||||||||||||||
Шеф-стюард | |||||||||||||||||
Заместитель шеф-стюарда | |||||||||||||||||
Ассистент технического делегата | |||||||||||||||||
Ассистент главного секретаря | 2 | ||||||||||||||||
Ассистент шеф-стюарда | |||||||||||||||||
Ассистент курс-дизайнера | |||||||||||||||||
Судья-стюард | |||||||||||||||||
Судья-секретарь | |||||||||||||||||
Судья-секундометрист | |||||||||||||||||
Читчик | |||||||||||||||||
Требования, обусловленные особенностями судейства | Для присвоения 1К необходимо в течение 2-х лет выполнить требования к прохождению практики судейства не менее 7-ми раз на соревнованиях соответствующего статуса в указанных должностях спортивного судьи | ||||||||||||||||
Требования для кандидатов, имеющих МСМК | 2 | ||||||||||||||||
1К присваивается МСМК по итогам прохождения семинара со сдачей квалификационного зачета на оценку не ниже «хорошо» и прохождения практики судейства на 2-х соревнованиях любого статуса в любых из следующих должностей: «ассистент курс-дизайнера», «судья-секретарь», «судья-секундометрист», «судья-стюард», «читчик». | |||||||||||||||||
Требования для кандидатов, имеющих МС | 3 | ||||||||||||||||
1К присваивается МС, имеющим 2К, но не ранее чем через 1 год со дня присвоения такой категории по итогам прохождения семинара со сдачей квалификационного зачета на оценку не ниже «хорошо» и прохождения практики судейства на 3-х соревнованиях соответствующего статуса в любых из следующих должностях спортивного судьи: «ассистент главного секретаря», «ассистент шеф-стюарда», «ассистент курс-дизайнера», «судья-стюард», «судья-секретарь», «судья-секундометрист», «читчик». | |||||||||||||||||
2К | 1 год | Ассистент главного секретаря | 5 | ||||||||||||||
Ассистент курс-дизайнера | |||||||||||||||||
Ассистент шеф-стюарда | |||||||||||||||||
Судья-секретарь | |||||||||||||||||
Судья-стюард | |||||||||||||||||
Судья-секундометрист | |||||||||||||||||
Читчик | |||||||||||||||||
Судья на старте/финише | |||||||||||||||||
Судья на препятствии/трассе | |||||||||||||||||
Требования, обусловленные особенностями судейства | Для присвоения 2К необходимо в течение 1-го года выполнить требования к прохождению практики судейства не менее 5-ти раз на соревнованиях соответствующего статуса в указанных должностях спортивного судьи | ||||||||||||||||
Требования для кандидатов, имеющих МС | 1 | ||||||||||||||||
2К присваивается МС, по итогам прохождения семинара со сдачей квалификационного зачета на оценку не ниже «хорошо» и прохождения практики судейства на одном соревновании любого статуса в любой из следующих должностей: «ассистент курс-дизайнера», «судья-секретарь», «судья-стюард», «судья-секундометрист», «читчик», «судья на препятствии/на трассе», «судья на старте/финише». | |||||||||||||||||
3К | Требования, обусловленные особенностями судейства | Для присвоения 3К необходимо выполнить требования к сдаче квалификационного зачета без выполнения требований к прохождению практики судейства | |||||||||||||||
ЮС | Требования, обусловленные особенностями судейства | Для присвоения ЮС необходимо выполнить требования к прохождению теоретической подготовки со сдачей квалификационного зачета без выполнения требований к прохождению практики судейства | |||||||||||||||
Общие требования, обусловленные особенностями судейства | 1. 2. Судейство международных спортивных соревнований, включенных в ЕКП, приравнивается к судейству всероссийских соревнований. 3. Под годом выполнения требований для присвоения квалификационной категории спортивного судьи понимается временной период в 12 месяцев, начинающийся с участия спортивного судьи в первом зачетном соревновании. | ||||||||||||||||
 Москвы, г. Санкт-Петербурга
Москвы, г. Санкт-Петербурга

1.2. Требования к прохождению практики судейства
для подтверждения квалификационных категорий
спортивных судей
Наименование квалификационной категории спортивного судьи | Период прохождения практики судейства | Наименования должностей спортивных судей, предусмотренных правилами вида спорта | Статус и наименование соревнований, их количество | ||||||||||||||
Чемпионат России | Кубок России | Первенство России, Всероссийская Спартакиада между субъектами Российской Федерации | Другие всероссийские спортивные соревнования, включенные в ЕКП | Чемпионат федерального округа, двух и более федеральных округов, чемпионаты г. | Первенство федерального округа, двух и более федеральных округов, первенства г. Москвы, г. Санкт-Петербурга | Другие межрегиональные соревнования, включенные в ЕКП | Чемпионат субъекта Российской Федерации (кроме г. Москвы и г. Санкт-Петербурга) | Кубок субъекта Российской Федерации | Первенство субъекта Российской Федерации (кроме г. Москвы и г. Санкт-Петербурга) | Другие официальные спортивные соревнования субъекта Российской Федерации | Чемпионат муниципального образования | Первенство муниципального образования | Другие официальные спортивные соревнования муниципального образования | ||||
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | |
ВК | 4 года | ГСК | Главный судья | 6 | |||||||||||||
Заместитель главного судьи | |||||||||||||||||
Главный секретарь | |||||||||||||||||
Заместитель главного секретаря | |||||||||||||||||
Курс-дизайнер | |||||||||||||||||
Шеф-стюард | |||||||||||||||||
Заместитель шеф-стюарда | |||||||||||||||||
Технический делегат | |||||||||||||||||
Судья-член Гранд-Жюри | |||||||||||||||||
Ассистент технического делегата | |||||||||||||||||
Ассистент главного секретаря | |||||||||||||||||
Ассистент шеф-стюарда | |||||||||||||||||
Ассистент курс-дизайнера | |||||||||||||||||
Требования, обусловленные особенностями судейства | Для подтверждения ВК необходимо в течение 4-х лет выполнить требования к прохождению практики судейства не менее 6 раз на соревнованиях соответствующего статуса в указанных должностях спортивного судьи, из которых не менее 1 всероссийского соревнования | ||||||||||||||||
Требования для допуска к судейству при неподтверждении ВК | 1 | 1 | |||||||||||||||
Судья допускается к судейству после выполнения требований к прохождению теоретической подготовки и требований к сдаче квалификационного зачета, включающего практическую часть (с оценкой не ниже «хорошо»). | |||||||||||||||||
1К | 2 года | ГСК | Главный судья | 5 | |||||||||||||
Заместитель главного судьи | |||||||||||||||||
Главный секретарь | |||||||||||||||||
Заместитель главного секретаря | |||||||||||||||||
Судья-член Гранд-Жюри | |||||||||||||||||
Технический делегат | |||||||||||||||||
Курс-дизайнер | |||||||||||||||||
Шеф-стюард | |||||||||||||||||
Заместитель шеф-стюарда | |||||||||||||||||
Ассистент технического делегата | |||||||||||||||||
Ассистент главного секретаря | |||||||||||||||||
Ассистент курс-дизайнера | |||||||||||||||||
Ассистент шеф-стюарда | |||||||||||||||||
Требования, обусловленные особенностями судейства | Для подтверждения 1К необходимо в течение 2-х лет выполнить требования к прохождению практики судейства не менее 5 раз на соревнованиях соответствующего статуса в указанных должностях спортивного судьи, из которых не менее 1-го раза на соревнованиях не ниже уровнем, чем «другие официальные спортивные соревнования субъекта Российской Федерации» | ||||||||||||||||
Требования для допуска к судейству при неподтверждении 1К | 1 | 1 | |||||||||||||||
Судья допускается к судейству после выполнения требований к прохождению теоретической подготовки и требования к сдаче квалификационного зачета, включающего практическую часть (с оценкой не ниже «хорошо»). | |||||||||||||||||
2К | 2 года | ГСК | Главный секретарь | ||||||||||||||
Заместитель главного секретаря | |||||||||||||||||
Судья-член Гранд-Жюри | |||||||||||||||||
Курс-дизайнер | |||||||||||||||||
Шеф-стюард | |||||||||||||||||
Заместитель шеф-стюарда | |||||||||||||||||
Ассистент тех. | |||||||||||||||||
Ассистент курс-дизайнера | |||||||||||||||||
Ассистент шеф-стюарда | |||||||||||||||||
Ассистент главного секретаря | |||||||||||||||||
Судья-секретарь | 4 | ||||||||||||||||
Судья-стюард | |||||||||||||||||
Судья-секундометрист | |||||||||||||||||
Читчик | |||||||||||||||||
Судья на старте/финише | |||||||||||||||||
Судья на препятствии/трассе | |||||||||||||||||
Требования, обусловленные особенностями судейства | Для подтверждения 2К необходимо в течение 2-х лет выполнить требования к прохождению практики судейства не менее 4 раз на соревнованиях соответствующего статуса в указанных должностях спортивного судьи | ||||||||||||||||
Требования для допуска к судейству при неподтверждении 2К | 1 | ||||||||||||||||
Судья допускается к судейству после выполнения требований к прохождению теоретической подготовки и требования к сдаче квалификационного зачета, включающего практическую часть (с оценкой не ниже «хорошо»). | |||||||||||||||||
3К | 1 год | Ассистент главного секретаря | |||||||||||||||
Ассистент курс-дизайнера | |||||||||||||||||
Ассистент курс-дизайнера | |||||||||||||||||
Ассистент шеф-стюарда | |||||||||||||||||
Судья-секретарь | 3 | ||||||||||||||||
Судья-стюард | |||||||||||||||||
Судья-секундометрист | |||||||||||||||||
Читчик | |||||||||||||||||
Судья на старте/финише | |||||||||||||||||
Судья на препятствии/трассе | |||||||||||||||||
Требования, обусловленные особенностями судейства | Для подтверждения 3К необходимо в течение 1-го года выполнить требования к прохождению практики судейства не менее 3 раз на соревнованиях соответствующего статуса в указанных должностях спортивного судьи | ||||||||||||||||
Требования для допуска к судейству при неподтверждении 3К | Необходимо выполнить требования к прохождению теоретической подготовки (участие в семинаре) и требования к сдаче квалификационного зачета (сдать квалификационный зачет) без выполнения требований к практике судейства | ||||||||||||||||
Общие требования, обусловленные особенностями судейства | 1. 2. Судейство международных спортивных соревнований, включенных в ЕКП, приравнивается к судейству всероссийских соревнований. 3. Под годом выполнения требований для подтверждения квалификационной категории понимается временной период в 12 месяцев, начинающийся со дня, следующего за датой приказа о присвоении (подтверждении) действующей квалификационной категории спортивного судьи. | ||||||||||||||||
 Москвы, г. Санкт-Петербурга
Москвы, г. Санкт-Петербурга Получить допуск к судейству в 1К. В течение 6 месяцев после сдачи квалификационного зачета необходимо пройти практику спортивного судейства на 2 соревнованиях указанного статуса в любых указанных выше судейских должностях. Подать в ОСФ ходатайство о подтверждении квалификационной категории с приложением копии карточки учета спортивной судейской деятельности.
Получить допуск к судейству в 1К. В течение 6 месяцев после сдачи квалификационного зачета необходимо пройти практику спортивного судейства на 2 соревнованиях указанного статуса в любых указанных выше судейских должностях. Подать в ОСФ ходатайство о подтверждении квалификационной категории с приложением копии карточки учета спортивной судейской деятельности. делегата
делегата Получить допуск к судейству в 3К. В течение 3 месяцев после сдачи квалификационного зачета необходимо пройти практику спортивного судейства на соревнованиях указанного статуса в любых указанных выше судейских должностях. Подать в РСФ ходатайство о подтверждении квалификационной категории с приложением копии карточки учета спортивной судейской деятельности.
Получить допуск к судейству в 3К. В течение 3 месяцев после сдачи квалификационного зачета необходимо пройти практику спортивного судейства на соревнованиях указанного статуса в любых указанных выше судейских должностях. Подать в РСФ ходатайство о подтверждении квалификационной категории с приложением копии карточки учета спортивной судейской деятельности. Засчитываются соревнования с оценкой качества судейства не ниже «хорошо».
Засчитываются соревнования с оценкой качества судейства не ниже «хорошо».1.3. Требования к оценке практики судейства
Наименование квалификационной категории спортивного судьи, проходящего практику судейства | Наименование квалификационной категории спортивного судьи, оценивающего практику судейства | Наименование должности спортивного судьи, оценивающего практику судейства, а также вид спорта, в соответствии с ВРВС, по которому такому спортивному судье присвоена квалификационная категория | Перечень нарушений (замечаний), выявленных в процессе судейства, влияющих на понижение оценок | Наименование и применяемая шкала оценок |
1 | 2 | 3 | 4 | 5 |
ВК | ВК, 1К | Работу членов ГСК, кроме главного судьи, оценивает главный судья/заместитель главного судьи. Работу ассистента главного секретаря и судей-секретарей оценивает главный секретарь/заместитель главного секретаря. Работу ассистента технического делегат оценивает технический делегат. Работу ассистента курс-дизайнера оценивает курс-дизайнер. | Безупречное исполнение судейских должностных обязанностей. Успешное и оперативное разрешение внезапно возникших ситуаций | Отлично |
1К | ВК, 1К, 2К | Исполнение стандартных судейских должностных обязанностей без ошибок | Хорошо | |
2К | ВК, 1К, 2К | Работу ассистента шеф-стюарда, судей-стюардов оценивает шеф-стюард/заместитель шеф-стюарда. Работу судьи на старте/финише, судьи на препятствии/трассе, читчика, судьи-секундометриста оценивает главный судья/заместитель главного судьи. Работу главного судьи оценивает ВКС или КС РСФ после рассмотрения официального отчета о соревнованиях в соответствии со статусом соревнований. Оценивать судейство имеет право только судья по виду спорта «конный спорт» | Исполнение судейских должностных обязанностей с некоторыми, не существенными, замечаниями или ошибками, не повлекшими искажения результатов, с нарушением графика работы, опозданиями | Удовлетворительно |
3К/ЮС | ВК, 1К, 2К | Исполнение должностных судейских обязанностей с существенными ошибками, повлекшими искажение результатов соревнований. Нарушение этических норм поведения спортивного судьи | Неудовлетворительно |
1. 4. Требования к прохождению теоретической подготовки
4. Требования к прохождению теоретической подготовки
Наименование присваиваемой (подтверждаемой) квалификационной категории спортивного судьи | Наименование квалификационной категории спортивного судьи, проходящего теоретическую подготовку | Наименование органа общероссийской или региональной спортивной федерации, или федерального органа, или подразделения федерального органа, ответственного за проведение теоретической подготовки | Наименование квалификационной категории спортивного судьи, ответственного за прохождение теоретической подготовки, а также вида спорта, в соответствии с ВРВС, по которому такому спортивному судье присвоена квалификационная категория | Количество теоретических занятий (академических часов) и форма их проведения (семинар, коллоквиум, круглый стол, иное) | |
В качестве участника | В качестве лектора | ||||
1 | 2 | 3 | 4 | 5 | 6 |
ВК <*> | ВК, претендующие на подтверждение категории, РК, ВК и 1К, претендующие на присвоение ВК | ВКС ОСФ | ВК по виду спорта «конный спорт» | Не менее 1 занятия в форме семинара в каждый год судейской деятельности. | Не менее 1 занятия в каждые 2 года судейской деятельности, в форме лекции или семинара лекция — не менее 3 часов, семинар — не менее 8 часов |
1К | 1К, претендующие на подтверждение категории, 2К, МСМК и МС, претендующие на присвоение 1К | КС РКФ | 1К, ВК по виду спорта «конный спорт» | Не менее 1 занятия в форме семинара в каждый год судейской деятельности. Для подтверждения — не менее 8 часов, для присвоения — не менее 16 часов | Не менее 1 занятия в каждые 2 года судейской деятельности, в форме лекции или семинара |
2К | 2К, претендующие на подтверждение категории, МС и 3К, претендующие на присвоение 2К | КС РКФ | 1К, ВК по виду спорта «конный спорт» | Не менее 1 занятия в форме семинара в каждый год судейской деятельности. | Не менее 1 занятия в каждые 2 года судейской деятельности, в форме лекции или семинара |
3К, ЮС | 3К, претендующие на подтверждение категории, кандидаты на присвоение 3К и ЮС | КС РКФ | 1К, ВК по виду спорта «конный спорт» | Не менее 1 занятия в форме семинара в каждый год судейской деятельности. Для присвоения — не менее 12 часов | |
<*> Участие судей, получивших или подтвердивших международную категорию по виду спорта «конный спорт», в международном семинаре по подготовке судей, засчитывается как выполнение требований к прохождению теоретической подготовки для присвоения (подтверждения) квалификационной категории спортивного судьи на текущий период. | |||||
 Для подтверждения — не менее 8 часов, для присвоения — не менее 16 часов
Для подтверждения — не менее 8 часов, для присвоения — не менее 16 часов Для подтверждения — не менее 7 часов, для присвоения — не менее 14 часов
Для подтверждения — не менее 7 часов, для присвоения — не менее 14 часов
1.6. Требования к сдаче квалификационного зачета (экзамена)
Наименование присваиваемой (подтверждаемой) квалификационной категории спортивного судьи | Требования для допуска к сдаче квалификационного зачета (экзамена) | Наименование квалификационной категории спортивного судьи, сдающего квалификационный зачет (экзамен) | Наименование квалификационной категории спортивного судьи, принимающего квалификационный зачет (экзамен), а также вид спорта, в соответствии с ВРВС, по которому спортивному судье присвоена соответствующая квалификационная категория | Наименование органа общероссийской или региональной спортивной федерации, ответственного за проведение квалификационного зачета (экзамена) и формирования тестовых вопросов (экзаменационных билетов) | Шкала оценок | Сроки и условия повторной сдачи квалификационного зачета (экзамена) | Периодичность сдачи квалификационного зачета (экзамена) (количество в год) |
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
ВК <*> | Выполнение требований к прохождению практики судейства с оценкой не ниже «хорошо» и теоретической подготовки, необходимых для присвоения/ подтверждения ВК | 1К, РК/ВС <**> (подтверждение — ВК) | ВК по виду спорта «конный спорт» | ВКС ОСФ | Зачет/Незачет. | Повторная сдача экзамена может быть разрешена не ранее, чем через три месяца | Не менее 1 раза в каждые 2 года судейской деятельности |
1К | Выполнение требований к прохождению практики судейства с оценкой не ниже «хорошо» и теоретической подготовки, необходимых для присвоения/ подтверждения 1К | 2К, МС, МСМК (подтверждение — 1К) | ВК, 1К по виду спорта «конный спорт» | КС РСФ | Зачет/Незачет. Теоретические и практические тесты должны содержать не менее 12 вопросов. | Повторная сдача экзамена может быть разрешена не ранее, чем через три месяца | Не менее 1 раза в каждые 2 года судейской деятельности |
2К | Выполнение требований к прохождению практики судейства с оценкой не ниже «хорошо» и теоретической подготовки, необходимых для присвоения/ подтверждения 2К | 3К, МС (подтверждение — 2К) | ВК, 1К по виду спорта «конный спорт» | КС РСФ | Зачет/Незачет. Теоретические и практические тесты должны содержать не менее 10 вопросов. Правильный ответ на один вопрос — 1 балл. Для получения оценки «Зачет» необходимо набрать не менее 70% от максимального количества баллов. | Повторная сдача экзамена может быть разрешена не ранее, чем через один месяц | Не менее 1 раза в каждый год судейской деятельности |
3К/ЮС | Выполнение требований к прохождению теоретической подготовки, необходимых для присвоения 3К/ЮС или требований к прохождению практики судейства и теоретической подготовки, необходимых для подтверждения 3К/ЮС | ЮС, кандидаты без категории (подтверждение — 3К/ЮС) | ВК, 1К по виду спорта «конный спорт» | КС РСФ | Зачет/Незачет. Теоретические и практические тесты должны содержать не менее 10 вопросов. Правильный ответ на один вопрос — 1 балл. Для получения оценки «Зачет» необходимо набрать не менее 60% от максимального количества баллов. | Повторная сдача экзамена может быть разрешена не ранее, чем через один месяц | Не менее 1 раза в каждый год судейской деятельности |
<*> Участие судей, успешно сдавших экзамен для получения или подтверждения МК в международном семинаре по подготовке судей, засчитывается как выполнение требований к сдаче квалификационного зачета для присвоения (подтверждения) ВК. <**> Для судей, подтверждающих категорию ВК, и имеющих практику судейства не менее 15 лет с категорией ВС/РК, допускается устное собеседование («круглый стол»). | |||||||
 Теоретические и практические тесты должны содержать не менее 15 вопросов. Правильный ответ на один вопрос — 1 балл. Для получения оценки «Зачет» необходимо набрать не менее 80% от максимального количества баллов <**>
Теоретические и практические тесты должны содержать не менее 15 вопросов. Правильный ответ на один вопрос — 1 балл. Для получения оценки «Зачет» необходимо набрать не менее 80% от максимального количества баллов <**> Правильный ответ на один вопрос — 1 балл. Для получения оценки «Зачет» необходимо набрать не менее 75% от максимального количества баллов.
Правильный ответ на один вопрос — 1 балл. Для получения оценки «Зачет» необходимо набрать не менее 75% от максимального количества баллов.

2. Требования к включению спортивных судей в судейские
коллегии
Наименования должностей спортивных судей, предусмотренных правилами спорта (располагаются по старшинству), их минимально допустимую квалификационную категорию и количество, с указанием функциональных обязанностей и подчиненности | Статус и наименование спортивных соревнований | ||||||||||||||
Чемпионат России | Кубок России | Первенство России, Всероссийская Спартакиада между субъектами Российской Федерации | Другие всероссийские спортивные соревнования, включенные в ЕКП | Чемпионат федерального округа, двух и более федеральных округов, чемпионаты г. | Первенство федерального округа, двух и более федеральных округов, чемпионаты г. Москвы, г. Санкт-Петербурга | Другие межрегиональные соревнования, включенные в ЕКП | Чемпионат субъекта Российской Федерации (кроме г. Москвы и г. Санкт-Петербурга) | Кубок субъекта Российской Федерации | Первенство субъекта Российской Федерации (кроме г. Москвы и г. Санкт-Петербурга) | Другие официальные спортивные соревнования субъекта Российской Федерации | Чемпионат муниципального образования | Первенство муниципального образования | Другие официальные спортивные соревнования муниципального образования | ||
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
Состав Главной судейской коллегии | |||||||||||||||
Главный судья | Квалификационная категория | ВК | 1К | ||||||||||||
Требования к возрасту | Не моложе 25 лет | Не моложе 23 лет | |||||||||||||
Количество | 1 | ||||||||||||||
Функциональные обязанности и подчиненность | Возглавляет судейскую коллегию, контролирует проведение соревнований, принимает решения в рамках своей юрисдикции. Главный судья имеет право делегировать часть своих полномочий своему заместителю, старшим судьям. | ||||||||||||||
Заместитель главного судьи | Квалификационная категория | ВК | 1К | ||||||||||||
Требования к возрасту | Не моложе 25 лет | Не моложе 23 лет | |||||||||||||
Количество | В зависимости от количества спортивных дисциплин. Один на каждую спортивную дисциплину. | ||||||||||||||
Функциональные обязанности и подчиненность | Назначается только в случае проведении турнира более чем по одной спортивной дисциплине. | ||||||||||||||
Главный секретарь | Квалификационная категория | ВК | 1К | 2К | |||||||||||
Требования к возрасту | Не моложе 25 лет | Не моложе 23 лет | Не моложе 21 года | ||||||||||||
Количество | 1 | ||||||||||||||
Функциональные обязанности и подчиненность | Руководит работой секретариата, отвечает за правильность ведения и оформления документации по соревнованиям. | ||||||||||||||
Заместитель главного секретаря | Квалификационная категория | ВК | 1К | 2К | |||||||||||
Требования к возрасту | Не моложе 25 лет | Не моложе 23 лет | Не моложе 21 года | ||||||||||||
Количество | В зависимости от количества спортивных дисциплин. | ||||||||||||||
Функциональные обязанности и подчиненность | Назначается только в случае проведении турнира более чем по одной спортивной дисциплине. Под контролем главного секретаря руководит работой секретариата в рамках делегированных ему главным секретарем полномочий: распределяет обязанности среди ассистентов главного секретаря, судей-секретарей и информаторов по своей спортивной дисциплине. Подчиняется главном судье, главному секретарю и заместителю главного судьи по своей спортивной дисциплине. | ||||||||||||||
Судья — член Гранд-Жюри | Квалификационная категория | 1К | 2К | ||||||||||||
Требования к возрасту | Не моложе 21 года | Не моложе 20 лет | Не моложе 18 лет | ||||||||||||
Количество | В спортивных дисциплинах, содержащих в своем наименовании «конкур» | ||||||||||||||
Не менее 2 | Не менее 1 | ||||||||||||||
В спортивных дисциплинах, содержащих в своем наименовании «выездка» | |||||||||||||||
Не менее 4 | Не менее 2 | ||||||||||||||
В спортивных дисциплинах, содержащих в своем наименовании «троеборье», «тентпеггинг» | |||||||||||||||
Не менее 2 | Не менее 1 | ||||||||||||||
В спортивных дисциплинах, содержащих в своем наименовании «драйвинг», «пробеги» | |||||||||||||||
Не менее 2 | |||||||||||||||
В спортивных дисциплинах, содержащих в своем наименовании «вольтижировка», «джигитовка» | |||||||||||||||
3 — 7 | 3 — 5 | ||||||||||||||
Условия | Количество судей — членов Гранд-Жюри увеличивается в соответствии с количеством спортивных дисциплин. Количество судей — членов Гранд-Жюри должно быть увеличено в зависимости от количества боевых полей. На каждое боевое поле должно приходиться судей — членов Гранд-Жюри (включая главного судью) — не менее количества, указанного выше. В спортивных дисциплинах, содержащих в своем наименовании «конкур» при количестве участников более 80 стартующих пар в день (на одном боевом поле) должен быть назначен как минимум один дополнительный судья. При наличии открытой канавы с водой должен быть назначен один дополнительный судья — член Гранд — жюри. В спортивных дисциплинах, содержащих в своем наименовании «выездка», «троеборье», «драйвинг» каждый судья — член Гранд-жюри (включая главного судью) имеет право судить не более 60 участников в день. | ||||||||||||||
Функциональные обязанности и подчиненность | Непосредственно оценивает выступление каждого участника, принимает решения в рамках своей юрисдикции. | ||||||||||||||
Технический делегат | Квалификационная категория | ВК | 1К | 1К/2К <*> | |||||||||||
Требования к возрасту | Не моложе 25 лет | ||||||||||||||
Количество | Не менее 1 | ||||||||||||||
Условия | На соревнованиях межрегионального уровня и выше в спортивных дисциплинах, содержащих в своем наименовании «троеборье» или «драйвинг» технический делегат не имеет права исполнять другие функции, кроме функций «технического делегата». Для соревнований регионального уровня и ниже в спортивных дисциплинах, содержащих в своем наименовании «троеборье» и «драйвинг» в порядке, установленном ОСФ, а также для соревнований любого уровня по другим спортивным дисциплинам функции технического делегата исполняет один из судей — членов Гранд-Жюри (кроме главного судьи). В случае проведения соревнований более чем по одной спортивной дисциплине технический делегат назначается на каждую спортивную дисциплину. <*> Для спортивных дисциплин, кроме спортивных дисциплин, содержащих в своем наименовании «троеборье», «драйвинг». | ||||||||||||||
Функциональные обязанности и подчиненность | Осуществляет надзор за технической стороной проведения соревнований, ведет анализ технических аспектов проведения соревнований и судейской работы, обладает правом трактовать Правила и Регламенты, следить за их исполнением, готовит отчет о соревнованиях. | ||||||||||||||
Курс-дизайнер | Квалификационная категория | ВК/1К <*> | 1К | 2К | |||||||||||
Требования к возрасту | Не моложе 25 лет | Не моложе 21 года | |||||||||||||
Количество | 1 — 2 | ||||||||||||||
Условия | Назначается на соревнования по спортивным дисциплинам, содержащих в своем наименовании «конкур», «троеборье», «драйвинг», «пробеги». <*> Для спортивных дисциплин «драйвинг» и «пробеги». | ||||||||||||||
Функциональные обязанности и подчиненность | Составляет схемы и дизайн маршрута/трассы каждого вида программы турнира; выставляет и измеряет маршрут конкура/паркура и/или трассу кросса/марафона/пробега, руководит работой ассистентов курс-дизайнера. Назначается из списка курс-дизайнеров, имеющих аттестацию на составление маршрутов/трасс соответствующей высоты/уровня сложности. Подчиняется главному судье соревнований. | ||||||||||||||
Шеф-стюард | Квалификационная категория | ВК | 1К | 2К | |||||||||||
Требования к возрасту | Не моложе 23 лет | Не моложе 21 года | |||||||||||||
Количество | 1 | ||||||||||||||
Функциональные обязанности и подчиненность | Организует стюардинг спортивных площадок, руководит работой заместителей шеф-стюарда, ассистентов шеф-стюарда и судей-стюардов, обеспечивает соблюдение кодекса благополучия по отношению к лошади и принципов «честной игры», принимает решения в рамках своей юрисдикции. | ||||||||||||||
Заместитель шеф-стюарда | Квалификационная категория | ВК | 1К | 2К | |||||||||||
Требования к возрасту | Не моложе 23 лет | Не моложе 21 года | |||||||||||||
Количество | В зависимости от количества спортивных дисциплин. Один на каждую спортивную дисциплину. | ||||||||||||||
Функциональные обязанности и подчиненность | Назначается только в случае проведении турнира более чем по одной спортивной дисциплине. | ||||||||||||||
Судьи, не входящие в состав Главной судейской коллегии | |||||||||||||||
Ассистент технического делегата | Квалификационная категория | 1К | 2К | ||||||||||||
Требования к возрасту | Не моложе 22 лет | ||||||||||||||
Количество | Не более 2 | ||||||||||||||
Условия | Может назначаться для спортивных дисциплин, содержащих в своем наименовании «троеборье» или «драйвинг» в порядке, установленном ОСФ. | ||||||||||||||
Функциональные обязанности и подчиненность | Оказывает помощь техническому делегату, выполняет возложенные на него техническим делегатом функции по контролю за техническими условия трасс кросса и марафона. Подчиняется техническому делегату соревнований. | ||||||||||||||
Ассистент главного секретаря | Квалификационная категория | 1К | 2К | 3К | |||||||||||
Требования к возрасту | Не моложе 21 года | Не моложе 19 лет | Не моложе 18 лет | ||||||||||||
Количество | 1 — 2 | Не более 1 | |||||||||||||
Функциональные обязанности и подчиненность | Оказывает помощь главному секретарю/заместителю главного секретаря, контролирует работу судей-секретарей в оформлении технической документации, связанной с проведением соревнований. | ||||||||||||||
Ассистент курс-дизайнера | Квалификационная категория | 1К | 2К | 3К | |||||||||||
Требования к возрасту | Не моложе 18 лет | Не моложе 16 лет | |||||||||||||
Количество | 1 — 4 | 1 — 2 | Не более 1 | ||||||||||||
Условия | Назначается на соревнования по спортивным дисциплинам, содержащих в своем наименовании «конкур», «троеборье», «драйвинг», «пробеги». | ||||||||||||||
Функциональные обязанности и подчиненность | Оказывает помощь курс-дизайнеру в разработке и постановке маршрута конкура/паркура и/или трассы кросса/марафона/пробега. Подчиняется курс-дизайнеру соревнований. | ||||||||||||||
Ассистент шеф-стюарда | Квалификационная категория | 1К | 2К | 3К | |||||||||||
Требования к возрасту | Не моложе 21 года | Не моложе 18 лет | |||||||||||||
Количество судей | 2 — 4 | 1 — 3 | Не более 2 | ||||||||||||
Функциональные обязанности и подчиненность | Оказывает помощь шеф-стюарду, контролирует работу судей-стюардов в своей зоне ответственности. | ||||||||||||||
Судья-секретарь | Квалификационная категория | 2К | 3К | 3К/ЮС | |||||||||||
Требования к возрасту | Не моложе 17 лет | Не моложе 16 лет | Не моложе 14 лет | ||||||||||||
Количество судей | 1 — 5 | Не более 3 | |||||||||||||
Функциональные обязанности и подчиненность | Регистрирует результаты выступления участников, в том числе с использованием специальных программ и/или оборудования. | ||||||||||||||
Судья-стюард | Квалификационная категория | 2К | 3К/ЮС | ||||||||||||
Требования к возрасту | Не моложе 17 лет | Не моложе 14 лет | |||||||||||||
Количество судей | 3 — 8 | 2 — 5 | Не более 3 | ||||||||||||
Функциональные обязанности и подчиненность | Осуществляет контроль за соблюдением Правил, Регламентов и кодекса благополучия лошади на территории проведения соревнований. | ||||||||||||||
Судья-секундометрист | Квалификационная категория | 2К | 3К | ||||||||||||
Требования к возрасту | Не моложе 16 лет | ||||||||||||||
Количество | Не более 8 | ||||||||||||||
Условия | Может назначаться на соревнования по всем спортивным дисциплинам, кроме содержащих в своем наименовании «выездка». | ||||||||||||||
Функциональные обязанности и подчиненность | Под руководством старшего судьи соревнования/главного судьи регистрирует время старта и/или финиша и/или время гита каждого участника, в том числе — с использованием специального электронного оборудования контроля и учета времени. | ||||||||||||||
Судья на старте/финише | Квалификационная категория | 3К/ЮС | |||||||||||||
Требования к возрасту | Не моложе 14 лет | ||||||||||||||
Количество | Не более 2 | ||||||||||||||
Условия | Может назначаться на соревнования по всем спортивным дисциплинам, кроме содержащих в своем наименовании «выездка», «вольтижировка». | ||||||||||||||
Функциональные обязанности и подчиненность | В случае недостаточной видимости линии старта/финиша судьями — членами ГСК и судьями-секундометристами обозначает пересечение каждым участником линии старта и/или финиша. | ||||||||||||||
Читчик | Квалификационная категория | 2К | 3К/ЮС | ||||||||||||
Требования к возрасту | Не моложе 17 лет | Не моложе 14 лет | |||||||||||||
Количество | 3 — 8 | Не более 8 | |||||||||||||
Условия | Может назначаться на соревнования по спортивным дисциплинам, содержащих в своем наименовании «выездка», «троеборье», «драйвинг», «вольтижировка», «джигитовка». | ||||||||||||||
Функциональные обязанности и подчиненность | Фиксирует оценки, выставленные судьей — членом Гранд-жюри каждого участника. | ||||||||||||||
Судья на препятствии/трассе | Квалификационная категория | 3К | |||||||||||||
Требования к возрасту | Не моложе 16 лет | ||||||||||||||
Количество | 2 — 20 | ||||||||||||||
Условия | Назначается на соревнования по спортивным дисциплинам, содержащих в своем наименовании «троеборье», «драйвинг», «пробеги». | ||||||||||||||
Функциональные обязанности и подчиненность | В спортивных дисциплинах, содержащих в своем наименовании «троеборье», «драйвинг» — определяет результат прохождения/преодоления препятствий участниками соревнований; в спортивных дисциплинах, содержащих в своем наименовании «драйвинг» — самостоятельно или с помощью судьи-секундометриста дополнительно фиксирует время прохождения препятствия каждым участником, в спортивных дисциплинах, содержащих в своем наименовании «троеборье», «драйвинг», «пробеги» — фиксирует прохождения контрольной точки каждым участником. | ||||||||||||||
1. Назначение судей производится с учетом аттестации ОСФ по спортивным дисциплинам и должностям спортивных судей. 2. Для соревнований более чем в одной спортивной дисциплине количество судей может быть увеличено в соответствии с количеством спортивных дисциплин. 3. Количество судей «не более» принимается в смысле: «должность является опциональной, количество не должно превышать указанного в данной строке», «не менее» — означает, что судья данной должности должен быть назначен, в количестве не меньшем, чем указано в данной строке. 4. Судейская коллегия всероссийских соревнований должна включать не менее 30% судей, представляющих иные субъекты Российской Федерации, чем регион, где проводятся соревнования. | |||||||||||||||
 Москвы, г. Санкт-Петербурга
Москвы, г. Санкт-Петербурга Назначает старшего судью каждого вида программы из числа судей — членов Гранд-жюри, распределяет обязанности среди судей — членов Гранд-Жюри.
Назначает старшего судью каждого вида программы из числа судей — членов Гранд-жюри, распределяет обязанности среди судей — членов Гранд-Жюри. Под контролем главного судьи руководит соревнованиями в рамках делегированных ему главным судьей полномочий: назначает старшего судью каждого вида программы по своей спортивной дисциплине, распределяет обязанности среди судей — членов Гранд-Жюри. Подчиняется главному судье соревнований.
Под контролем главного судьи руководит соревнованиями в рамках делегированных ему главным судьей полномочий: назначает старшего судью каждого вида программы по своей спортивной дисциплине, распределяет обязанности среди судей — членов Гранд-Жюри. Подчиняется главному судье соревнований. Контролирует работу ассистентов главного секретаря, судей-секретарей, информатора, в рамках своей юрисдикции отвечает за публикацию информации о ходе соревнований для участников и зрителей. Главный секретарь имеет право делегировать часть своих полномочий своему заместителю. Подчиняется главному судье соревнований.
Контролирует работу ассистентов главного секретаря, судей-секретарей, информатора, в рамках своей юрисдикции отвечает за публикацию информации о ходе соревнований для участников и зрителей. Главный секретарь имеет право делегировать часть своих полномочий своему заместителю. Подчиняется главному судье соревнований. Один на каждую спортивную дисциплину.
Один на каждую спортивную дисциплину.
 В течение соревнований должна быть обеспечена ротация судей — членов Гранд-Жюри в разных программах. В соответствии с Правилами из числа судей — членов Гранд-Жюри в каждом виде программы назначается старший судья. В спортивных дисциплинах, кроме спортивных дисциплин, содержащих в своем наименовании слова «драйвинг» и «троеборье», также один из судей-членов Гранд-Жюри дополнительно выполняет функции технического делегата. Подчиняется главному судье/заместителю главного судьи.
В течение соревнований должна быть обеспечена ротация судей — членов Гранд-Жюри в разных программах. В соответствии с Правилами из числа судей — членов Гранд-Жюри в каждом виде программы назначается старший судья. В спортивных дисциплинах, кроме спортивных дисциплин, содержащих в своем наименовании слова «драйвинг» и «троеборье», также один из судей-членов Гранд-Жюри дополнительно выполняет функции технического делегата. Подчиняется главному судье/заместителю главного судьи.
 В спортивных дисциплинах, содержащих в своем наименовании «драйвинг» и «троеборье», также осуществляет контроль над техническими условия и подготовкой трасс марафона и кросса. Подчиняется главному судье соревнований.
В спортивных дисциплинах, содержащих в своем наименовании «драйвинг» и «троеборье», также осуществляет контроль над техническими условия и подготовкой трасс марафона и кросса. Подчиняется главному судье соревнований. В соревнованиях по спортивным дисциплинам, содержащих в своем наименовании «троеборье», «драйвинг» как минимум один курс-дизайнер должен иметь аттестацию на постановку кросса/марафона.
В соревнованиях по спортивным дисциплинам, содержащих в своем наименовании «троеборье», «драйвинг» как минимум один курс-дизайнер должен иметь аттестацию на постановку кросса/марафона. Подчиняется главному судье соревнований.
Подчиняется главному судье соревнований. Под контролем шеф-стюарда руководит работой службы стюардинга по своей спортивной дисциплине. Руководит работой ассистентов шеф-стюарда и судьями-стюардами по своей спортивной дисциплине. Подчиняется шеф-стюарду.
Под контролем шеф-стюарда руководит работой службы стюардинга по своей спортивной дисциплине. Руководит работой ассистентов шеф-стюарда и судьями-стюардами по своей спортивной дисциплине. Подчиняется шеф-стюарду.
 Подчиняется главному секретарю/заместителю главного секретаря.
Подчиняется главному секретарю/заместителю главного секретаря.
 Подчиняется шеф-стюарду и/или заместителю шеф-стюарда.
Подчиняется шеф-стюарду и/или заместителю шеф-стюарда. Выполняет функции, возложенные на него главным секретарем и/или заместителем главного секретаря. Подчиняется главному секретарю и/или заместителю главного секретаря.
Выполняет функции, возложенные на него главным секретарем и/или заместителем главного секретаря. Подчиняется главному секретарю и/или заместителю главного секретаря. Выполняет функции, возложенные на него шеф-стюардом и/или ассистентом шеф-стюарда. Подчиняется шеф-стюарду, заместителю шеф-стюарда, ассистенту шеф/стюарда.
Выполняет функции, возложенные на него шеф-стюардом и/или ассистентом шеф-стюарда. Подчиняется шеф-стюарду, заместителю шеф-стюарда, ассистенту шеф/стюарда. Подчиняется судьям — членам Гранд-Жюри.
Подчиняется судьям — членам Гранд-Жюри.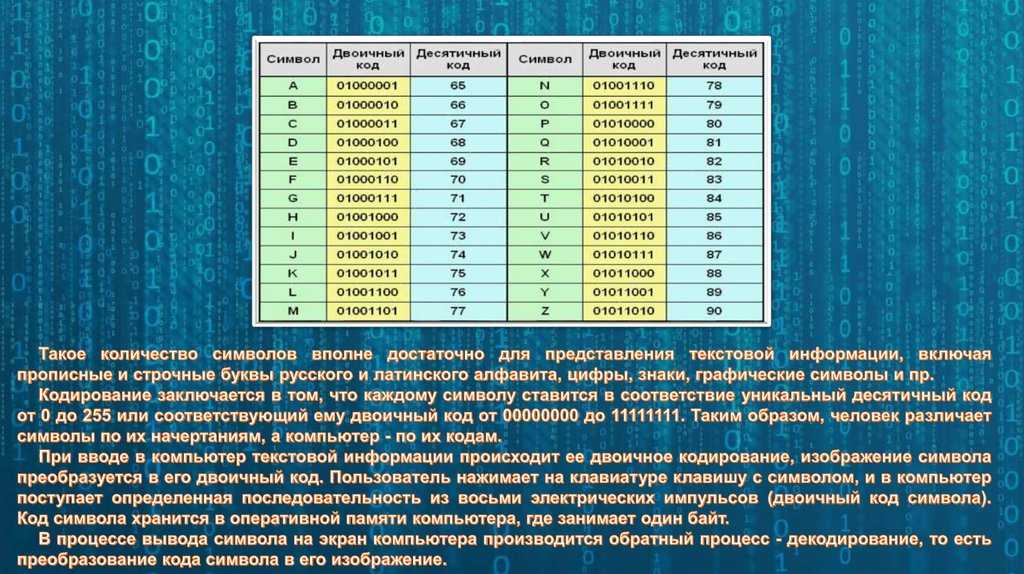 Подчиняется судьям — членам Гранд-Жюри.
Подчиняется судьям — членам Гранд-Жюри. В спортивных дисциплинах, содержащих в своем наименовании «выездка», «вольтижировка», «джигитовка» читчик старшего судьи дополнительно контролирует правильность исполнения и время выполнения схемы теста участником. Подчиняется судьям — членам Гранд-Жюри.
В спортивных дисциплинах, содержащих в своем наименовании «выездка», «вольтижировка», «джигитовка» читчик старшего судьи дополнительно контролирует правильность исполнения и время выполнения схемы теста участником. Подчиняется судьям — членам Гранд-Жюри. Подчиняется главному судье соревнований и/или заместителю главного судьи по соответствующей спортивной дисциплине и техническому делегату.
Подчиняется главному судье соревнований и/или заместителю главного судьи по соответствующей спортивной дисциплине и техническому делегату.Сокращения, используемые в настоящих квалификационных
требованиях к спортивным судьям по виду спорта
«конный спорт»
ВК — квалификационная категория спортивного судьи «спортивный судья всероссийской категории»;
1К — квалификационная категория спортивного судьи «спортивный судья первой категории»;
2К — квалификационная категория спортивного судьи «спортивный судья второй категории»;
3К — квалификационная категория спортивного судьи «спортивный судья третьей категории»;
ЮС — квалификационная категория спортивного судьи «юный спортивный судья»;
РК — квалификационная категория спортивного судьи «судья по спорту республиканской категории»;
ВС — квалификационная категория спортивного судьи «судья по спорту всесоюзной категории»;
ОСФ — общероссийская спортивная федерация по виду спорта «конный спорт»;
РСФ — региональная спортивная федерация по виду спорта «конный спорт»;
ГСК — главная судейская коллегия;
КС — коллегия судей:
ВКС — всероссийская коллегия судей;
МС — мастер спорта России по виду спорта «конный спорт»;
МСМК — мастер спорта России международного класса по виду спорта «конный спорт»;
ЕКП — Единый календарный план межрегиональных, всероссийских и международных физкультурных мероприятий и спортивных мероприятий;
Правила — правила вида спорта «конный спорт»;
ВРВС — Всероссийский реестр видов спорта;
Положение — Положение о проведении официального спортивного соревнования или физкультурного мероприятия;
МК — спортивный судья международной категории;
Соревнование — официальное спортивное соревнование или физкультурное мероприятие, проводимое в соответствии с правилами вида спорта.
Понимание соотношения сторон в фотографии
Давайте подробно рассмотрим соотношения сторон фото вместе с профессиональным фотографом Назимом Мансуровым и выясним, что нужно знать каждому обо всем этом.
Многие современные смартфоны имеют нативное соотношение сторон 4:3 на получаемых изображениях. Изображение снято на iPhone X @ 4mm, ISO 40, 1/15, f/1.8
Что такое соотношение сторон?
В фотографии соотношение сторон представляет из себя соотношение между шириной и высотой изображения.
Общие пропорции в фотографии
Соотношение сторон часто определяется форм-фактором датчика пленки/изображения камеры, который практически всегда является прямоугольным. Наиболее распространенные пропорции современных сенсоров цифровых камер — 3:2 и 4:3. Все современные полнокадровые и зеркальные фотокамеры APS-C имеют сенсоры с соотношением сторон 3:2, тогда как 4:3 — популярный выбор среди смартфонов, Micro Four Thirds и некоторых производителей камер среднего формата.
Некоторые камеры позволяют выбирать различные соотношения сторон в меню камеры, предоставляя опции обрезки, отличные от оригинального датчика изображения.
Когда соотношение сторон выражается двумя числами, разделенными двоеточием, первое число обычно относится к горизонтальной стороне изображения, а второе число — к вертикальной. Например, 3:2 обозначает горизонтальное изображение, полученное в альбомной ориентации. А 2:3 будет считаться вертикальным изображением, полученным в портретной. Когда соотношение сторон выражается в десятичных числах, таких как 1,50 или 1,50:1, оно игнорирует ориентацию изображения.
Почему соотношение сторон важно
CMOS-датчики Nikon имеют соотношение сторон 3:2
Понимание основ соотношения сторон очень важно, потому что это влияет на окончательное изображение. Это может быть особенно важно во время физического захвата фотографии. Например, если вы снимаете изображение с помощью камеры с его собственным соотношением сторон 4:3 и втискиваете свой объект или важные элементы сцены в края кадра, вы не сможете обрезать изображение с более широкими пропорциями. Посмотрите на следующее изображение.
Посмотрите на следующее изображение.
Как вы можете видеть, изображение было получено в 4:3, и фотограф едва смог втиснуть здание, а также структуру переднего плана в кадр. Хотя в конце этого конкретного снимка это сработало, просто нет возможности обрезать изображение, чтобы оно соответствовало любым другим форматным соотношениям, без обрезания в здание или элемент переднего плана.
То же самое касается выбора чрезвычайно широких форматов изображения, когда камера обрезает верх и низ кадра, как на фото ниже:
Это изображение было получено в формате 16:9 Каппадокия, Турция. Снято с помощью DJI Mavic Pro @ 10,26 мм, ISO 100, 1/60, f/5,6
Когда Назим использовал беспилотник DJI Mavic Pro в Каппадокии, Турция, он забыл, что настроил камеру для съемки изображений в формате 16:9, поэтому получил кучу широких изображений, подобных фото, которое вы видите выше. К сожалению, поскольку верхняя и нижняя части изображения были обрезаны (даже при съемке в формате RAW), пришлось немного обрезать края изображения, чтобы получить 3:2 или 4:3. Посмотрите на то, что изменение этих пропорций сделает с изображением выше.
Посмотрите на то, что изменение этих пропорций сделает с изображением выше.
То же изображение, показывающее, что происходит во время кадрирования с соотношением сторон 3:2 и 4:3
Как вы видите 4:3 — определенно неудачный вариант, поскольку он врезается в элементы переднего плана. Если бы фотограф запечатлел это изображение в его нативном формате 4:3 для начала, он мог бы избежать этой проблемы.
Вот почему очень важно обращать внимание на пропорции при составлении композиции и кадрировании снимков — всегда обеспечивайте достаточное «пространство для дыхания» вокруг объекта съемки, если ваша конечная цель состоит в том, чтобы позже было больше вариантов кадрирования.
Нативное соотношение против выбора в камере
Соотношение сторон часто определяется датчиком изображения камеры, которое является «родным» для нее. Однако некоторые камеры предоставляют фотографам возможность выбирать разные пропорции.
Например, Nikon Z7 позволяет выбирать между следующим:
- FX (36×24)
- DX (24×16)
- 5:4 (30×24)
- 1:1 (24×24)
- 16:9 (36×20)
Обратите внимание, что первые два параметра имеют формат 3:2 (FX 36×24 и DX 24×16), поскольку это нативное соотношение сторон сенсора на этой камере (второй вариант предназначен для обрезки центральной части изображения для моделирования датчиков камеры Nikon APS-C/DX). Все остальные параметры, такие как 5:4, 1:1 и 16:9, не являются встроенными, что означает, что выбор любого из них приведет к обрезке части изображения.
Все остальные параметры, такие как 5:4, 1:1 и 16:9, не являются встроенными, что означает, что выбор любого из них приведет к обрезке части изображения.
Собор Святой Софии, запечатленный в оригинальном соотношении сторон камеры 3:2 NIKON Z 7 + NIKKOR Z 24-70 мм f/4 S @ 70 мм, ISO 64, 1/6, f/5,6
Хотя это уменьшает разрешение изображения и размер файла, часто не стоит переключаться на невстроенные пропорции. Прежде всего, вы выбрасываете пиксели, которые никогда не сможете вернуть. Если вы обрежете изображение и решите вернуться и изменить его на другое соотношение сторон, вам придется либо повторно сделать снимок, либо потенциально потерять разрешение из-за дополнительного кадрирования. Если вы снимаете в исходном соотношении сторон, у вас будет возможность изменить его на этапе последующей обработки с минимальной потерей разрешения.
Почему кто-то может захотеть изменить исходное соотношение сторон? Первая причина связана с кадрированием — если вы хотите предотвратить случайную обрезку объекта, то может быть полезно переключиться на соотношение сторон, которое вы будете использовать для отображения или печати изображения. Вторая причина связана с буфером камеры и скоростью непрерывной съемки — некоторые камеры могут снимать дольше из-за хранения меньших файлов в буфере.
Вторая причина связана с буфером камеры и скоростью непрерывной съемки — некоторые камеры могут снимать дольше из-за хранения меньших файлов в буфере.
Полное солнечное затмение, снято в формате 4:3 Fujifilm GFX 50S + GF23mmF4 R LM WR @ 23 мм, ISO 100, 10 сек, f/11,0
Если вы хотите максимизировать разрешение изображений, всегда снимайте в исходном соотношении сторон датчика камеры. Однако если у вас есть особые требования (например, ваш клиент просит, чтобы вы производили изображения с определенным соотношением сторон), то кадрирование в камере будет более безопасным вариантом с точки зрения композиции.
Наиболее распространенные пропорции
Ниже приведены наиболее распространенные пропорции
- 1:1 (1,00) — Некоторые пленочные камеры среднего формата предлагают соотношение сторон 1:1. Тем не менее, ни одна из современных цифровых камер не имеет квадратных датчиков, и только некоторые камеры предлагают 1:1 в качестве опции в меню. Instagram сделал популярным 1:1, изначально применяя его к каждой фотографии, но платформа была изменена для соответствия различным форматам изображения. 1:1 является относительно распространенным для печати квадратных изображений.
- 5:4 (1,25). Соотношение сторон 5:4 довольно распространено в пленочных камерах большого и среднего формата, а также при печати изображений размером 8 ″ x10 ″ и 16 ″ x20 ″.
- 3:2 (1,50). Большинство зеркальных, беззеркальных и компактных камер имеют сенсоры 3:2, независимо от их размера. Соотношение сторон 3:2 было популяризировано 35-мм пленкой, и сегодня оно является наиболее распространенным в фотографии.
- 4:3 (1,33) — средний формат, Micro Four Thirds, большинство смартфонов и некоторые камеры типа «наведи и снимай» имеют сенсоры 4:3.
- 16:9 (1,78) — самый распространенный формат видео сегодня. Не распространенный формат в фотографии, но некоторые камеры предоставляют его в качестве опции кадрирования.
- 3:1 (3,0) — обычно используется для панорамных снимков.
 1:1 является относительно распространенным для печати квадратных изображений.
1:1 является относительно распространенным для печати квадратных изображений.Панорама Дед-Хорс-Поинт в штате Юта с примерным соотношением сторон 3:1 Hasselblad X1D @ 45 мм, ISO 100, 1/125, f/5,6
Лучшие в мире гонки на вертикальные километры
|
Все мнения являются нашими собственными и не зависят от брендов. Если вы покупаете по ссылкам на этой странице, мы можем получить небольшую комиссию. Вот наш процесс.
Забеги по вертикальным километрам — распространенная концепция гонок в европейском горном беге, но я не слышал о них, пока не начал исследовать и посещать Альпы для трейлового бега.
РЕКЛАМА
Итак, что такое вертикальный километр…?
«горный бег в гору, который проходит примерно 1000 метров, преодолевая дистанцию не более 5 км» часто упоминается как «ВК» или «ВКМ».
В то время как широкое определение указывает, что дистанция обычно не превышает 5 км, многие гонки на вертикальные километры короче, а некоторые иногда длиннее (например, Трансвулкания). Вертикальный километр Шамони, который мы пробежали (но не участвовали в гонках), составляет менее 4 км.
Нам нравится концепция «пробега» вертикального километра, так как она настолько проста: максимальные усилия столько времени, сколько потребуется, чтобы подняться на 1000 м по горной местности — и вы будете вознаграждены фантастическим видом с вершины горы.
Восхождение по маршруту «Вертикальный километр Шамони».В некоторых из этих гонок есть «массовые старты», когда все начинают гонку одновременно. В других гонках VK, таких как Dolomites VK, небольшие группы людей начинают волны, а в других, например, в Шамони, один бегун стартует каждые 30 секунд. У каждого типа старта есть свои преимущества, с намерением, чтобы один человек стартовал за раз, что в основном избегает заторов на узких однопутных трассах (хотя вам все равно может потребоваться обгонять или быть обогнанным бегунами вокруг вас в какой-то момент).
Некоторые из этих гонок начинаются почти на уровне моря, а другие, например Санта-Катерина ВК, начинаются на относительно большой высоте. Учитывая сочетание максимальных усилий, необходимых для непрерывного лазания, и увеличения высоты, будьте готовы к сверхтяжелой работе, особенно если вы живете и тренируетесь ближе к уровню моря!
Broken Arrow SkyraceВертикальные километры обычно проводятся в то же время, что и одна или несколько других, более длинных гонок, включая гонки Skyrunning и горные ультрамарафоны.
- Пятница, день/ранний вечер: Проходит гонка «Вертикальный километр»
- Суббота: Начало ультрамарафонской гонки (ультрасы часто начинаются в субботу, так как их прохождение может занять много часов!)
- Воскресенье: Проходит гонка Skyrace Это означает, что если ваши ноги/тело справятся с этим, вы можете посетить удивительный горный городок, пробежать Вертикальный километр в пятницу и еще одну гонку в эти выходные – наиболее вероятным вариантом будет Skyrace (я не думаю, что многие люди участвуют в гонках ВКонтакте в один день, а на следующее утро пробежать ультрамарафон, но, возможно, некоторые так и делают…).
Гонки на вертикальный километр в Великобритании
В Великобритании есть несколько великолепных гор, но не так много таких, на которые можно подняться на полную 1000 м за 5 км или меньше. Событие Glen Coe Skyrace является исключением и является единственной гонкой на вертикальный километр в Великобритании: их мероприятие VK, Mamores VK , охватывает 1000 м на 5 км.
СВЯЗАННЫЕ: Бег по пересеченной местности с палками: преимущества, недостатки и советы
Европейские забеги на вертикальные километры
В европейских горах, родине скайраннинга и вертикальных километров, вы избалованы выбором. Вот подборка тех, которые волнуют нас больше всего!
- Chamonix, France (1000 м больше 3,8 км)
- Доломиты, Италия (1000 м более 2,6 км)
- , Швейцария (1000 м больше 1,9 км — Стипт! , Греция (1024 м на 4,3 км)
- Трансвулкания, Канарские острова, Испания (на 1200 м с набором высоты 7,6 км эта гонка длиннее, чем «традиционное» определение гонки «Вертикальный километр» — еще более сложная задача!)
- Тромсё, Норвегия (1044 м на дистанции 2,7 км)
- Зегама-Айзкорри, Испания (1015 м на дистанции 3 км на время (с 2,2-километровым участком без учета времени)
- Сантана, Мадейра, Португалия 0
(10083 км) на дистанции- Санта-Катерина, Италия (1000 м на 2,9 км)
Североамериканские гонки на вертикальные километры
По мере того, как горный бег и гонки в европейском стиле становятся все более популярными, мы увидим все больше и больше американских и канадских гонок на вертикальные километры.
- Флагстафф, Аризона (1000 м на 5 км… включая несколько спусков, чтобы вы могли накопить достаточное вертикальное усиление на этой дистанции)
- Лоун Пик, Монтана (1106 м на 5 км, в гоночный уик-энд «The Rut»)
- Уайтфейс-Маунтин, Нью-Йорк (1000 м на 4 км)
- Broken Arrow Skyrace VK, Скво-Вэлли, Калифорния (945 м на 5 км — не совсем ВК, но заслуживает упоминания за свою эпичность)
СВЯЗАННЫЕ: Broken Arrow Skyrace: отчет о гонке, фотографии и советы Есть выбор: 700 м на 3 км или полные 1000 м на 5 км.
Не останавливайтесь на 900 метрах — вы прошли 90% пути!Сколько времени у меня займет вертикальный километр?
Если вы являетесь обладателем мирового рекорда по вертикальному километру, Kilian Jornet , или другой супер-хардкорный горный бегун, то вы сможете пробежать ВК менее чем за 40 минут! Нынешними (по состоянию на 2017 год) мировыми рекордсменами среди мужчин и женщин являются 90 009 Урбан Земмер (29 м 42 с!) и 90 009 Кристель Девалле (34 м 44 с) 90 010 соответственно, которые оба установили это время на самой крутой трассе в Фулли, Швейцария, в 2014 году.
Для простых смертных прохождение курса по-прежнему является большим достижением, и это может занять у вас пару часов в зависимости от курса, условий, а также вашего опыта и уровня физической подготовки.
Хотя это будет болезненно, независимо от того, сколько времени это займет, вас будут поддерживать чувство достижения и эпические виды, которые вы сможете получить в качестве награды!
Мы всегда рады узнавать о новых авантюрных маршрутах и гонках. Дайте нам знать в комментариях, если мы пропустили какие-либо замечательные гонки на вертикальные километры из нашего списка, и расскажите нам немного о них!
СВЯЗАННЫЕ СООБЩЕНИЯ
ТРЕНДЫ НА T&K
РЕКЛАМА
ПОСЛЕДНИЕ О T&K
Вертикальные гонки на километры | Советы по тренировкам
Мэтт Мейнард участвует в забеге
© iancorless.com
Последние 15 лет или около того стали свидетелями всплеска популярности гонок на вертикальный километр — практики бега в гору как можно быстрее.
Введенная в этом году в рамках серии UK Skyrunner® гонка Salomon Mamores VK™ в Глен-Коу, Шотландия, является первым британским мероприятием VK. Мэтт Мейнард принял участие в гонке в эти выходные (16-18 сентября). Вот его советы о том, как тренироваться для этого.
ЕЩЕ: Узнайте, каково это пробежать 3000 м по вертикали
Salomon Mamores Участник VK™ Франсуа Гонон
© iancorless.com
ровная дорога работает, чтобы преуспеть в гонке ВК. Но вам не нужно переезжать в Альпы, чтобы тренироваться. Просто выберите местный холм и бегите вверх по нему. Спринты должны варьироваться по продолжительности от 8-15 секунд. В конце каждого повторения медленно возвращайтесь назад, прежде чем приступить к следующему усилию.
Это должна быть тренировка максимальной интенсивности, но я всегда нахожу ужасным вставать с постели в День подъема. Если у вас проблемы с мотивацией, лучше провести тренировку с меньшей интенсивностью или с меньшим количеством повторений, чем оставить шторы закрытыми и нетронутыми кроссовки.
Вертикальный по названию, вертикальный по своей природе
© guillemcasanova.com
2. Свет ступней
Когда Эдмунд Хиллари поднялся на Эверест в 1953 году, он сказал знаменитую фразу: «Один фунт на ногах равен пяти фунтам на спине». .» С тех пор это непропорциональное соотношение подтвердилось в различных исследованиях.
Я выбрал обувь Inov-8 Talon 212. При весе (как вы уже догадались) 212 г его легкость была идеальной. Его мягкая резина также любит грызть крутую травянистую местность на трассе Salomon Mamores VK ™, а верхняя часть обуви прочна и устойчива к разрывам в области носка, где многие другие бренды находят свой конец.
Трасса самая крутая в средней части
© Harvey Maps
3. Изучите маршрут
Salomon Mamores VK™ поднимается на 1000 м на своем 5-километровом маршруте, но некоторые подъемы VK набирают эту высоту всего за 1,92 км! Знание маршрута означает, что вы сможете мысленно разбить задачу.
Были участки, где я знал, что буду царапать руками. В тот день, однако, самой нервной частью был ровный спринт на 600 м на старте мимо толпы!
Кто сказал, что нельзя участвовать в гонках в килте?
© guillemcasanova.com
4. Подготовьтесь
Если вы можете пробежать 10 км по ровной поверхности за час, то прохождение маршрута ВКонтакте займет у вас два-три часа. Уровень усилий также похож на 10-километровую гонку. Вы должны быть готовы работать почти на максимальной частоте сердечных сокращений, достигая такой одышки, которая означает, что вам будет трудно сказать «привет» болельщикам. (Мировой рекорд для ВКонтакте — 29 минут и 42 секунды.)
Несколько испанских друзей были рядом с вершиной и подбадривали меня — я так старался, что не смог сказать «Привет!»
Вид с вершины на Кинлохлевен
© iancorless.
5. Правильно питайтесь
Последний прием пищи вам нужно будет сделать за два-три часа до гонки. Не пейте воду в последний час. Поэкспериментируйте с энергетическим гелем с кофеином непосредственно перед тренировками по спринту в гору. Если ваш желудок хорошо реагирует, съешьте его непосредственно перед соревнованиями. Я использовал гель с кофеином SIS, потому что его легче глотать и он немного менее вязкий, чем другие бренды.
Короткие шаги помогают экономить энергию
© iancorless.com
6. Сделайте глубокий вдох
Salomon Mamores VK™ — это гонка на время, в которой участники начинают каждые 30 секунд, мчась на время к вершине. Выполните высокоинтенсивную разминку, которая заставит вас тяжело дышать. За одну минуту до выхода на посадку сделайте глубокий, успокаивающий вдох и убедите себя, что все ваши тренировки вот-вот окупятся.
Последний скалистый спринт к вершине
© iancorless.com
7.
Первый участок маршрута представляет собой 600 м ровной быстрой дороги. После этого вы окажетесь на подъеме, но по проходимой тропе. Делайте короткие шаги, чтобы экономить энергию.
Часто во время гонок по холмистой местности меня обгоняли идущие люди. На этот раз я знал лучше, и примерно на полпути пришло время увеличить мощность.
ВКонтакте — это не обычная гонка
© iancorless.com
8. Мощный поход
Текущий рекордсмен ВКонтакте Урбан Земмер редко бегает во время соревнований. Исследования показали, что на склонах с крутизной более 28% спортсмены могут снизить расход энергии при ходьбе.
Перенесите свой вес на бедра и сильно надавите на ведущую ногу. На некоторых беговых шортах до бедра теперь нанесена цепкая резиновая печать поверх материала. Идея состоит в том, что вы можете сильнее нажимать на ноги без соскальзывания рук и, таким образом, передавать больше силы через руки в ноги. Некоторые участники ВКонтакте также используют палки для ходьбы.
Или, что еще лучше, отправляйтесь в горы и потренируйтесь в силовом походе!
Болельщики собираются на гребне вершины
© iancorless.com
9. Отдайте все, что у вас есть
Большинство гонок ВКонтакте заканчиваются на вершине горы — например, На Гругайчан на Salomon Mamores VK™. В течение последних пяти минут гонки вам больше не нужно беспокоиться о своем темпе — разгоняйтесь, пока не задохнетесь, гарантируя, что вы закончите без остатка в баке. После восхождения быстро укутаться теплой одеждой. Перекусывайте по мере продвижения вниз и не забывайте подбадривать своих товарищей-конкурентов.
Участник Франсуа Гонон втягивает воздух
© guillemcasanova.com
10. Принеси колокольчик
Эта гонка может быть первой в своем роде в Великобритании, но ВКонтакте пользуется популярностью на континенте, особенно во Франции. , годами. На таких гонках, как Chamonix, Partisan Tour De France, толпы людей собираются в горах и звонят в колокольчики, чтобы выразить свою поддержку.
БОЛЬШЕ: 5 лучших прибрежных походов в Корнуолле
Для получения дополнительной информации посетите сайт skylinescotland.com/mamores-vertical-kilometre.
Фото: iancorless.org, guillemcasanova.com
Мэтт завершил ВК на 33-м месте из 160 участников со временем 00:56:19. Следите за ним в Твиттере.
Инсерция ДНК путем вырезания и вставки с использованием модифицированных CRISPR-ассоциированных транспозаз типа V-K
Новые результаты
Просмотреть профиль ORCIDConnor J. Tou, Просмотреть профиль ORCIDBenno Orr, Просмотреть профиль ORCIDBenjamin P. Kleinstiver
DOI: https://doi.org/10.1101/2022.01.07.475005
-2 [PubMed] [CrossRef] [Google Scholar]
- Аннотация
- Полный текст
- Информация/история
- Metrics
- Difement
- Metrics
- .
Main
Программируемая вставка последовательностей ДНК с несколькими тысячами оснований в геномы без использования гомологичной рекомбинации и двухцепочечных разрывов (DSB) открывает новые возможности для точного редактирования генома. Методы геномной интеграции обычно основаны на вирусных векторах 1,2 или транспозонах 3-7 , оба из которых лишены программируемости и поэтому встраиваются в геном стохастически, или нуклеазах, связанных с донорами ДНК 8-10 , которые основаны на цитотоксичности.
Два основных класса CAST, типы I и V-K, обладают различными и взаимодополняющими свойствами. В то время как охарактеризованные CAST типа I проявляют высокую специфичность к мишени и обычно приводят только к предполагаемым продуктам простого вставочного гена 17 (хотя и с исключениями 18 ), большее количество генов Cas, стехиометрическая сложность и размер кодирования могут ограничивать дальнейшее развитие инструментов в других организмах, таких как эукариотические клетки.
Рисунок 1. Разработка и характеристика HELIX.
a — c , Схемы CAST типа I и типа V-K и HELIX (панели a — c соответственно) и их механизмы транспозиции, которые приводят к простой вставке или коинтеграции генных продуктов. d , Рабочий процесс для экспериментов по транспозиции, нацеленных на плазмидные субстраты. e , Транспозиция оценивалась с помощью ПЦР соединения через LE/RE в TS1 в pTarget. Эксперименты проводили с nAniI, слитым с N- или С-концом TnsB, при использовании pDonor без сайтов I-AniI. f , Количественная оценка эффективности интеграции ДНК с плазмидами при использовании ShHELIX и донорной плазмиды с диапазоном расстояний (d) между сайтом I-AniI и LE/RE, оцененным с помощью ddPCR с использованием минипрепарированной ДНК. g , Покрытие ожидаемых продуктов вставки в pTarget из секвенирования с длинным чтением с использованием подмножества примерных простых чтений вставки для ShHELIX и чтений коинтеграции для ShCAST (покрытие из чтений коинтеграции ShHELIX и чтений простого вставки ShCAST опущено для простоты) .
CAST типа I и типа V-K различаются по наличию или отсутствию субъединицы TnsA, соответственно, одно из основных отличий, определяющее их простое введение для коинтеграции чистоты продукта. Как в транспозонах Tn7, так и в CAST типа I, TnsA и TnsB выполняют реакции разрыва 5” и 3” доноров соответственно, что приводит к простым вставкам посредством транспозиции вырезания и вставки ( Fig. 1a ). В транспозонах Tn5053 и CAST типа V-K, лишенных TnsA, а также в транспозонах Tn7 и модифицированных системах I типа с каталитически мертвым TnsA 17,20 , только 3-дюймовый донорский надрез происходит через TnsB. Доноры с одиночным надрезом приводят к значительной доле коинтегратных вставок за счет репликативной транспозиции вместо вырезания и вставки 21 ( Fig. 1b ). Таким образом, для создания коинтегративного CAST типа V-K мы предположили, что восстановление отсутствующей функции TnsA путем слияния ортогональной ДНК-никазы с TnsB позволит транспозицию «вырезать и вставить» через 5-дюймовые и 3-дюймовые донорные надрезы (, рис.
Одним из семейств ферментов, соответствующих этим критериям, являются самонаводящиеся эндонуклеазы (HEs), которые представляют собой небольшие нуклеазы, способные генерировать DSB с относительно высокой специфичностью.
Результаты
Разработка и оптимизация HELIX
Сначала мы определили, может ли nAniI адекватно заменить отсутствие TnsA в транспозоне V-K канонического типа из Scytonema hofmannii (ShCAST) 14 . Для этого мы сконструировали серию экспрессионных плазмид ShHELIX, каждая из которых содержала: (1) одну направляющую РНК (sgRNA), нацеленную на сайт-мишень 1 (TS1) на отдельной плазмиде-мишени (pTarget), (2) Cas12k, (3) TniQ, (4) TnsC и (5) nAniI, слитые с N- или C-концом TnsB ( Рис. 1d ). Экспрессионные плазмиды ShCAST или ShHELIX котрансформировали донорной плазмидой (pDonor, содержащей груз размером 2,1 т.п.н.) с левым и правым концами транспозона (LE и RE соответственно) в штамм E. coli , несущий pTarget (, рис. 1d). ). Чтобы определить, произошла ли транспозиция при использовании слияний nAniI с TnsB, мы провели ПЦР соединения как LE, так и RE в pTarget на минипрепарированной ДНК из соскобленных колоний.
Затем мы собрали восемь дополнительных донорных плазмид, несущих переменные расстояния между LE/RE и сайтом-мишенью I-AniI (ориентированных на создание 5-дюймового никеля на pDonor) ( рис. 1f ). При совместной трансформации экспрессионных плазмид ShCAST или ShHELIX (с N-концевым слиянием nAniI-TnsB) вместе с pDonor в наш штамм pTarget мы наблюдали одинаковое количество трансформантных колоний при использовании конструкций ShCAST и ShHELIX, что указывает на отсутствие дефекта жизнеспособности клеток из-за HELIX. ( доп. рис. 1а ). Мы оценили эффективность интеграции с помощью капельной цифровой ПЦР (ddPCR), а с помощью ShHELIX мы наблюдали диапазон эффективности интеграции при различных интервалах на pDonor, при этом интервал 14 п.
Затем мы использовали секвенирование с длительным чтением, чтобы оценить, может ли восстановление 5-дюймового никеля на pDonor с помощью ShHELIX улучшить чистоту продукта по сравнению с каноническим ShCAST. Мы обогатили транспонированные продукты из нашего пула минипрепарированных плазмид (, суп. рис. 2, ), линеаризовали обогащенную плазмидную ДНК и выполнили секвенирование с длительным считыванием, чтобы определить долю простых вставок в коинтегратах ().0009 Рис. 1г-1и ). С помощью ShCAST мы наблюдали 19,39% коинтеграций, что согласуется с предыдущими результатами 6 ( рис. 1i ). Поразительно, что ShHELIX почти полностью устранил коинтеграты, что привело к сокращению всего продукта до 0,6% (уменьшение в 32 раза по сравнению с ShCAST; рис. 1h и 1i ).
Мы также провели серию контрольных экспериментов для получения дополнительных характеристик ShHELIX ( Sup. Note 2 ). Во-первых, каталитически аттенуированный вариант I-AniI (K227M, Q171K) уменьшал коинтеграцию в 1,8 раза по сравнению с ShCAST (предположительно из-за неполной инактивации надрезов I-AniI) ( Sup. Fig. 4a ). Во-вторых, pDonor, лишенный сайта-мишени I-AniI, приводил к снижению коинтегратов в 1,7 раза по сравнению с ShCAST ( Sup. Fig. 4a ). Затем эксперименты с использованием pDonor с «перевернутым» сайтом I-AniI, который размещает разрыв на той же цепи, что и разрыв TnsB, привели к 10-кратному уменьшению коинтегратов ( Sup.
Характеристика HELIX на геномных мишенях
Воодушевленные нашими результатами транспозиции на плазмидных мишенях, мы затем исследовали эффективность ShHELIX-опосредованной интеграции ДНК в геномные сайты. Мы выполнили трансформации с использованием конструкций, сходных с экспериментами по нацеливанию плазмид, но вместо этого с sgRNAs, нацеленными на геном, и без pTarget (, рис. 2a, ). Во-первых, мы проверили влияние двух аминокислотных линкеров разной длины между nAniI и TnsB на эффективность геномной интеграции в нашем наборе из восьми донорных плазмид, содержащих различные расстояния между сайтами I-AniI и LE/RE. Эксперименты проводили с ранее охарактеризованной sgRNA 9.0324 14 против геномного сайта-мишени (TS2). Для обоих аминокислотных линкеров мы наблюдали самую высокую эффективность интеграции при расстоянии 14 п.н. между сайтом I-AniI и LE/RE (, рис. 2b, ), что согласуется с нашими результатами нацеливания плазмиды. Все обнаруживаемые вставки были в ориентации T-LR (, рис.
Рисунок 2. Характеристика вставок ДНК на геномных мишенях с помощью HELIX.
a , Рабочий процесс для экспериментов по транспозиции генома. b , Эффективность интеграции при использовании двух разных аминокислотных линкеров между nAniI и TnsB, sgRNA против геномного сайта-мишени 2 (TS2) и набора из восьми донорных плазмид с различными расстояниями между сайтами I-AniI и LE/RE , как определено с помощью ddPCR. c , Процент ориентации вставки при использовании ShCAST или ShHELIX, нацеленного на TS2, и с использованием pDonor с интервалом 14 п. панель) и относительную интеграцию с ShHELIX, нормализованную по ShCAST (правая панель), оцененную с помощью ddPCR. e , Охват ожидаемых продуктов вставки в геном (TS2) в результате секвенирования с длинным чтением с использованием подмножества типичных простых чтений вставки для ShHELIX и чтений коинтеграции для ShCAST (покрытие из чтений коинтеграции ShHELIX и чтений простых вставок ShCAST опущено для простоты).
Определив оптимальный сайт I-AniI для расстояния между LE/RE на pDonor для нацеливания на геном, мы затем сравнили эффективность интеграции и чистоту продукта ShCAST и ShHELIX для ряда геномных сайтов. ShHELIX сохранил надежную целевую интеграцию под управлением РНК в шести геномных сайтах-мишенях на уровнях, сравнимых с ShCAST (, рис. 2d, ). Чтобы проанализировать чистоту продукта интеграции HELIX при нацеливании генома на TS2, мы выполнили секвенирование с длительным считыванием с использованием стратегии целевого обогащения Cas9 32 . Анализ прочтений, обогащенных мишенью, при использовании ShCAST и ShHELIX, которые содержали или не содержали вставку груза, показал, что эффективность интеграции, рассчитанная на основе наших данных секвенирования с длинным чтением, была аналогична нашим результатам ddPCR на TS2 ( Sup. Fig. 5 ). С помощью ShCAST мы наблюдали, что 46,52% прочтений вставок были коинтегрированными ( рис. 2e-g ), что в целом ниже, чем наблюдалось ранее, хотя и в отношении другого целевого сайта и с помощью альтернативных методов секвенирования длинных прочтений 17 .
Затем мы оценили способность ShHELIX интегрировать ДНК-грузы различных размеров. Мы выполнили трансформации, используя донорные плазмиды, несущие грузы последовательности 5,2, 7,8 или 9,8 т.п.н. (по сравнению с pDonor с грузом 2,1 т.п.н., использованным в предыдущих экспериментах). Для каждой реакции транспозиции с использованием более крупных грузов ShHELIX показал сравнительно высокую эффективность направленной интеграции ДНК независимо от размера груза (9).0009 Рис. 2h ). В совокупности наши результаты демонстрируют, что ShHELIX способен к высокоактивным, однонаправленным вставкам ДНК путем вырезания и вставки и нечувствителен к грузам размером не менее 10 т.п.н.
Минимизация архитектуры ShCAST и HELIX
В отличие от многокомпонентной сложности CAST типа I, для которой требуется семь или восемь белковых субъединиц различной стехиометрии, мы задались вопросом, можем ли мы еще больше упростить обычно 4-субъединичную архитектуру CAST типа V-K.
Рис. 3. Эффективность интеграции трехкомпонентных систем CAST и HELIX.
a , Схема архитектур слияния для создания 3-х компонентных ШКАСТов. b , Эффективность интеграции, нормализованная к канонической, 4-компонентной системе ShCAST для каждой 3-компонентной системы ShCAST, как определено с помощью ddPCR (см. Sup. Fig. 6a для абсолютных уровней интеграции). c , Схема 3-х компонентных систем ШХЕЛИКС. d , Эффективность интеграции, нормализованная для 4-компонентной системы ShHELIX для каждой 3-компонентной системы ShHELIX (см. Sup. Fig. 6b для абсолютных уровней интеграции). Для панелей b и d среднее значение, стандартное отклонение и отдельные точки данных показаны для n = 3,9.0005
Затем мы определили, были ли аналогичные слияния Cas12k аналогичными функциональными в контексте 3-компонентных систем HELIX (, рис.
Расширяемость HELIX для ортологов CAST типа V-K
Во всех обнаруженных CAST типа V-K отсутствует TnsA 36 .
Рис. 4. Расширение HELIX для ортологов CAST типа V-K.
a , Филогенетическое дерево, иллюстрирующее разнообразие последовательностей TnsB из недавно идентифицированных CAST типа V-K 36 CAST, использованных в настоящем исследовании, а также Tn5053, отмечены. b , конструкции sgRNA для AcCAST. c , Эффективность интеграции с AcCAST с использованием двух конструкций sgRNA (из панели b ) и донорной плазмиды с нативной фланкирующей последовательностью (как сообщалось ранее 14 ) или фланкирующей последовательности ShCAST, оцененной с помощью ddPCR.
Чтобы исследовать применимость HELIX к другим ортологам CAST, мы охарактеризовали и оптимизировали два ранее опубликованных CAST типа V-K либо из Anabaena cylindrica (AcCAST), либо из другого штамма Scytonema hofmannii (ShoCAST).
Мы сконструировали AcHELIX, включающий слияние nAniI-TnsB вместе с конструкцией sgRNA-2 и pDonor, содержащим сайты I-AniI в 14 п.н. от LE/RE, разделенные фланкирующей последовательностью ShCAST (, рис. 4d, ). Чтобы определить чистоту продукта интеграции с AcHELIX по сравнению с AcCAST при нацеливании на геном, мы выполнили секвенирование с длительным считыванием после Cas9.обогащение мишени ( рис. 4e ). В то время как с AcCAST мы наблюдали 37,83% продуктов коинтеграции, для AcHELIX мы обнаружили только 0,71%, что представляет собой 53-кратное улучшение чистоты продукта с AcHELIX (, рис.
Затем мы охарактеризовали ShoCAST и ShoHELIX с использованием pDonor с интервалом 14 п.н., разделяющим сайт I-AniI, и LE/RE с фланкирующей последовательностью ShCAST (, рис. 4k, ). Мы провели эксперименты по нацеливанию генома с помощью ShoCAST и ShoHELIX, используя ранее описанную sgRNA 9.0324 16 против TS2.
Обсуждение
CAST — новый класс технологий редактирования генома, которые позволяют программировать вставки ДНК, не полагаясь на рекомбинацию, специфичные для последовательности рекомбиназы или DSB.
Мы демонстрируем, что HELIX эффективен для трех ортологов CAST типа V-K, подтверждая универсальность этого подхода. Продолжающееся открытие CAST не только позволит выявить новые системы с полезными характеристиками, но также создаст платформу для разработки технологий HELIX следующего поколения с разнообразными и полезными свойствами. Продолжающееся метагеномное открытие CAST выявило множество архитектур транспозонов с рядом свойств, включая компактные CAST типа V-K, которые могут оказаться полезными для приложений редактирования генома 36 . Оптимизация свойств HELIX для каждого ортолога CAST (например, аминокислотные линкеры, промежутки между сайтами nHE и LE/RE, выбор nHE, фланкирующие донорные последовательности и т. д.) приведет к более высокой эффективности транспозиции при одновременном использовании уникальных свойств каждого CAST.
Среди нашей характеристики CAST мы также обнаружили, что фланкирующее секвенирование, непосредственно примыкающее к LE/RE на pDonor, может влиять на эффективность интеграции. Это открытие предполагает, что будущие усилия по лучшему пониманию и увеличению фланкирующих последовательностей и потенциально других параметров CAST обещают дальнейшее повышение эффективности транспозиции. Более полное понимание факторов, определяющих эффективность интеграции CAST и HELIX и чистоту продукта (например, фланкирующие последовательности донора, выбор целевого сайта, размер груза, более полное определение PAM и т. д.), а также специфичность будут иметь решающее значение при разработке и настройка этих систем.
Подводя итоги, можно сказать, что HELIX представляет собой обобщающий подход, который обеспечивает программируемые, вырезающие и вставляемые, однонаправленные вставки ДНК с минимальным количеством компонентов и независимые от рекомбинации без DSB. Усилия по созданию высокоспецифичных и гиперактивных вариантов CAST типа V-K, потенциально с помощью инженерии TnsC и Cas12k или TnsB и Cas12k соответственно, могут принести пользу системам HELIX следующего поколения.
Вклады
C.J.T задумал идею, спроектировал и провел эксперименты, а также написал черновой вариант рукописи. Б.О. провел эксперименты для ShoHELIX и сравнения размеров груза. Б.П.К. участвовал в планировании эксперимента и руководил исследованием. CJT и B.P.K. отредактировали рукопись; все авторы одобрили рукопись.
Конкурирующие интересы
C.J.T. и B.P.K являются изобретателями патентов и/или патентных заявок, поданных Mass General Brigham, которые описывают технологии геномной инженерии. Б.П.К. является консультантом Avectas Inc., EcoR1 Capital и ElevateBio, а также советником Acrigen Biosciences, Life Edit Therapeutics и Prime Medicines.
Доступность данных
Все данные секвенирования будут храниться в архиве NCBI Sequence Read Archive (SRA).
Методы
Плазмиды и олигонуклеотиды
Все плазмиды, использованные в этом исследовании, и выбранные последовательности перечислены в Дополнительной таблице 1 . Новые плазмиды были созданы с помощью изотермической сборки или сборки Golden Gate, некоторые из которых были депонированы с помощью Addgene ( Supplementary Table 1 ). Плазмиды pHelper и pDonor для ShCAST и AcCAST, а также pTarget были подарены Feng Zhang (номер плазмиды Addgene 1279).21, 127924, 127923, 127925, 127926). Для плазмид, кодирующих гРНК, спейсерные последовательности клонировали в плазмиды pCAST и pHELIX посредством сборки Golden Gate с SapI (New England Biolabs, NEB). Особенности сайта-мишени для всех гРНК, использованных в этом исследовании, приведены в дополнительной таблице 2 . Олигонуклеотиды и зонды, используемые в этом исследовании, были приобретены у Integrated DNA Technologies (IDT) и перечислены в дополнительной таблице 3 .
Реакции транспозиции, направленные на плазмиды и геномные сайты
Трансформации для экспериментов по нацеливанию плазмид проводили в химически компетентных клетках PIR1, содержащих pTarget (исходный штамм PIR1, полученный от Invitrogen), с использованием 25 нг pCAST или pHELIX и 25 нг pDonor. Трансформированные клетки восстанавливали в течение 1 часа при 37°C в S.O.C. и затем высевали на чашки с агаром LB, содержащие 50 мкг/мл канамицина, 25 мкг/мл хлорамфеникола и 100 мкг/мл карбенициллина. Планшеты инкубировали при 37°С в течение 18 часов. Колонии подсчитывали, соскребали и экстрагировали плазмидную ДНК с помощью минипрепарата (Qiagen). Полученный пул плазмид использовали для последующего анализа с помощью соединительной ПЦР и секвенирования с длинным считыванием. ПЦР соединения анализировали с помощью капиллярного электрофореза QIAxcel (Qiagen) и визуализировали с помощью программного обеспечения QIAxcel ScreenGel (v1.
Трансформации для экспериментов по нацеливанию генома выполняли с использованием клеток PIR1 (или клеток PIR2 (Invitrogen) для Sup. Fig. 7 ) и 25 нг pCAST или pHELIX и 25 нг pDonor. Трансформированные клетки восстанавливали в течение 1 часа при 37°C в S.O.C. и затем высевали на чашки с агаром LB, содержащие 50 мкг/мл канамицина и 100 мкг/мл карбенициллина. Для трансформаций, включающих плазмиды ShCAST, ShHELIX, ShoCAST или ShoHELIX, планшеты инкубировали при 37°C в течение 18 часов; для трансформаций AcCAST и AcHELIX планшеты инкубировали при 37 °C в течение 24 часов из-за сравнительно меньших колоний (хотя их количество было приблизительно одинаковым). Колонии соскребали и гДНК собирали с использованием набора для очистки геномной ДНК Wizard (Promega) для последующего анализа с помощью ddPCR и секвенирования с длительным считыванием.
Оценка эффективности интеграции с помощью ddPCR
Плазмидную или геномную ДНК сначала нормализовали до 10 нг/мкл или 100 нг/мкл соответственно, а затем дополнительно разбавляли до 0,2 нг/мкл или 2 нг/мкл рабочих растворов соответственно для анализы ддПЦР.
Длинное секвенирование плазмид и геномных интеграций
Чистоту продукта интеграции анализировали с помощью длительного секвенирования с использованием плазмид, полученных в результате реакций транспозиции плазмид.
Для проведения длительного секвенирования целевых вставок генома мы выполнили протокол целевого обогащения Cas9 без амплификации для селективного улучшения секвенирования предполагаемых сайтов-мишеней (Oxford Nanopore Technologies, SQK-CS9109; sgRNA, перечисленные в дополнительной таблице ).
Обработка данных долгосрочных результатов секвенирования
Файлы Fast5 вызывались в режиме реального времени с помощью Miknow (v21.06.9) с моделью быстрого базового вызова, а полученные файлы FastQ были отфильтрованы для оценки Q > 8. BBDuk из Пакет BBTools 37 использовали для фильтрации ридов, содержащих 20 п.н. LE, RE и спейсерную последовательность целевого сайта с максимальным расстоянием Хэмминга 2. ), обнаруженные в остове плазмиды (не ожидается, что они появятся в продуктах простой вставки), были классифицированы как потенциальные коинтеграты, а те, которые не содержат эту последовательность, были классифицированы как потенциальные простые вставки. Чтения для экспериментов по нацеливанию на плазмиду дополнительно отфильтровывали для соответствующей длины чтения. Чтения, содержащие продукты, назначенные как простые вставки или коинтеграции, были объединены в один файл FastQ и сопоставлены либо с синтетической простой вставкой, либо с продуктом коинтеграции с Minimap2 38 указан с параметром map-ont .
Анализ расстояния вставки с использованием целевого секвенирования
Расстояние вставки от PAM до LE оценивали с помощью секвенирования следующего поколения с использованием метода конструирования библиотеки на основе двухэтапной ПЦР. 50 нг геномной ДНК из экспериментов по нацеливанию на геном подвергали ПЦР-амплификации с 30 циклами с использованием высокоточной ДНК-полимеразы Q5 (NEB) и праймеров, которые связываются сразу за пределами TS2 или сразу внутри LE. Продукты ПЦР очищали с использованием парамагнитных шариков, приготовленных, как описано ранее 9.0324 39,40 . 20 нг очищенного продукта ПЦР использовали в качестве матрицы для второго 10-циклового ПЦР для добавления штрих-кодов Illumina и последовательностей адаптеров.
Обработка данных целевых результатов секвенирования
Парные чтения FastQ сначала были отфильтрованы для Q>30 с помощью BBDuk из набора BBTools и объединены с помощью BBMerge. Затем были извлечены чтения, содержащие 20 п.н. TS2 и 20 п.н. терминального LE, каждое с максимальным расстоянием Хэмминга 1. Каждое чтение затем удаляли из последовательности выше и включая PAM и ниже и включая LE, в результате чего оставалась только последовательность между PAM и LE (т.е. сайт вставки). Длины результирующих прочтений были рассчитаны и использованы для построения профилей расстояния вставки PAM-LE.
Благодарности
Мы благодарим Л.
Ссылки
- 1.↵
Hendrie, P.C. and Russell, D.W. Нацеливание генов с помощью вирусных векторов. Молекулярная терапия 12, 9–17 (2005).
- 2.↵
Томас, С. Э., Эрхардт, А. и Кей, М. А. Прогресс и проблемы с использованием вирусных векторов для генной терапии. Nature Reviews Genetics 4, 346–358 (2003).
- 3.↵
Tellier, M., et al. Маринер и суперсемейство транспозонов ITm. Микробиолог. Спектр . 3, MDNA3–0033–2014 (2015)
- 4.↵
van Opijnen, T. and Camilli, A. Секвенирование вставок транспозонов: новый инструмент для системного анализа микроорганизмов.
- 5.
Ханифорд, Д. Б. и Эллис, М. Дж. Транспозоны Tn10 и Tn5. Микробиолог. Спектр . 3, MDNA3–0002–2014 (2015).
- 6.↵
Пластерк, Р. Х. А., Изсвак, З. и Ивич, З. Инопланетяне-резиденты: надсемейство мобильных элементов TC1/моряк. Trends in Genetics 15, 326–332 (1999).
- 7.↵
Wilson, M.H., Coates, C.J. and George, A.L. Транспозон-опосредованный перенос генов Piggybac в клетках человека. Молекулярная терапия 15, 139–145 (2007).
- 8.↵
Мали, П. и др. РНК-управляемая инженерия генома человека с помощью cas9. Наука 339, 823–826 (2013).
- 9.↵
Cong, L. et al. Мультиплексная инженерия генома с использованием систем CRISPR/Cas. Наука 339, 819–823 (2013).
- 10.↵
Rouet, P., Smih, F.
- 11.↵
Wang, H.H. et al. Программирование клеток с помощью мультиплексной инженерии генома и ускоренной эволюции. Природа 460, 894–898 (2009).
- 12.↵
Wang, H.H. et al. Разработка промотора в масштабе генома путем совместной селекции MAGE. Нац. методы 9, 591–593 (2012).
- 13.↵
Jiang, W., et al. Редактирование бактериальных геномов с помощью РНК с использованием систем CRISPR-Cas. Нац. Биотехнолог . 31, 233–239 (2013).
- 14.↵
Strecker, J. et al. Вставка ДНК под управлением РНК с CRISPR-ассоциированными транспозазами. Наука 365, 48–53 (2019).
- 15.↵
Klompe, S.E. et al. Системы CRISPR-CAS, кодируемые транспозонами, направляют интеграцию ДНК под управлением РНК.
- 16.↵
Vo, P. L. et al. CRISPR РНК-управляемые интегразы для высокоэффективной мультиплексной инженерии бактериального генома. Nature Biotechnology 39, 480–489 (2020).
- 17.↵
Vo, PL, Acree, C., Smith, M.L. & Sternberg, SH. Беспристрастное профилирование продуктов транспозиции, управляемой РНК CRISPR, путем секвенирования с длинным считыванием. Мобильная ДНК 12, (2021).
- 18.↵
Saito, M. et al. Двойной режим CRISPR-ассоциированного самонаведения транспозонов. Сотовый 184, (2021).
- 19.↵
Strecker, J. et al. Ответ на комментарий к «Вставка ДНК под управлением РНК с CRISPR-ассоциированными транспозазами». Наука 368, (2020).
- 20.↵
Мэй, Э. В. и Крейг, Н. Л. Переход от вырезания и вставки к репликативной транспозиции TN7. Наука 272, 401–404 (1996).
- 21.↵
Холодий Г.Ю. и др. Четыре гена, два конца и область res участвуют в транспозиции TN5053: парадигма для нового семейства транспозонов, несущих мероперон или интегрон. Молекулярная микробиология 17, 1189–1200 (1995).
- 22.↵
Сюй, С.-юн. Эндонуклеазы, специфичные для последовательности ДНК. Биомолекулярные концепции 6, 253–267 (2015).
- 23.↵
Гасюнас Г. и др. Рибонуклеопротеиновый комплекс Cas9-crРНК опосредует специфическое расщепление ДНК для адаптивного иммунитета у бактерий. Труды Национальной академии наук 109, (2012).
- 24.↵
Jinek, M. et al. Программируемая ДНК-эндонуклеаза с двойной РНК в адаптивном бактериальном иммунитете. Наука 337, 816–821 (2012).
- 25.↵
Hickman, A.B. et al. Неожиданное структурное разнообразие в рекомбинации ДНК. Molecular Cell 5, 1025–1034 (2000).
- 26.↵
Сюй, С. и Гупта, Ю.К. Природные эндонуклеазы HNH ленты цинка и инженерная эндонуклеаза разрезания цинковых пальцев. Nucleic Acids Research 41, 378–390 (2012).
- 27.↵
Стоддард, Б.Л. Самонаводящиеся эндонуклеазы из мобильных интронов группы I: от открытия до геномной инженерии. Мобильная ДНК 5, 7 (2014).
- 28.↵
McConnell Smith, A. et al. Генерация никирующего фермента, который стимулирует сайт-специфическую конверсию генов из самонаводящейся эндонуклеазы I-ani LAGLIDADG. Труды Национальной академии наук 106, 5099–5104 (2009).
- 29.↵
Takeuchi, R., et al. Оптимизация in vivo активности бифункциональной самонаводящейся эндонуклеазы и матуразы обращает вспять эволюционную деградацию. Nucleic Acids Research 37, 877–890 (2008).
- 30.↵
Querques, I., et al. Выбор и ремоделирование целевого сайта с помощью систем CRISPR-Transposon типа V.
- 31.↵
Скалли-Ким, М., МакКоннелл-Смит, А. и Стоддард, Б.Л. Коэволюция самонаводящейся эндонуклеазы и ее целевой последовательности-хозяина. Журнал молекулярной биологии 372, 1305–1319 (2007).
- 32.↵
Гилпатрик, Т. и др. Направленное секвенирование нанопор с лигированием адаптера под контролем cas9. Nature Biotechnology 38, 433–438 (2020).
- 33.
Парк, Ю.Ю. и другие. Структурная основа для выбора сайта-мишени в системах транспозиции ДНК, управляемых РНК. Наука 373, 768–774 (2021).
- 34.
Чой, К.Ю., Спенсер, Дж.М. и Крейг, Н.Л. Регулятор транспозиции TN7 TnsC взаимодействует с субъединицей транспозазы tnsb и селектором мишени TNSD. Труды Национальной академии наук 111, (2014).
- 35.
Mizuno, N. et al. Mub представляет собой AAA+ АТФазу, которая образует спиральные филаменты для контроля выбора мишени для транспозиции ДНК.
- 36.↵
Rybarski, J. R., et al. Метагеномное открытие CRISPR-ассоциированных транспозонов. Труды Национальной академии наук 118(49) e2112279118 (2021).
- 37.↵
BBMap – Bushnell B. – sourceforge.net/projects/bbmap/
- 38.↵
нуклеотидных последовательностей для парных выравниваний. Биоинформатика 34, 3094–3100 (2018).
- 39.↵
Роланд, Н. и Райх, Д. Экономичные высокопроизводительные библиотеки секвенирования ДНК для мультиплексного захвата мишеней. Рез. генома . 22, 939–946 (2012).
- 40.↵
Kleinstiver, B.P., et al. Разработаны варианты CRISPR-Cas12a с повышенной активностью и расширенными диапазонами нацеливания для генного, эпигенетического и базового редактирования. Нац. Биотехнолог . 37, 276–282 (2019).
Наверх
VK Treadmill Challenge — Trail Running Factory
Автор: Андрюс Рамонас Тренер Trail Running Factory
Удивительно, но за более чем 20 лет бега я ни разу не пытался серьезно тренироваться в горах на беговой дорожке. Чтобы не бегать под проливным дождем, однажды днем я пробежал ВК (1000 метров вертикального подъема) на дредмиле (…беговой дорожке). Поскольку беговая дорожка автоматически останавливается через 60 минут, я решил, что часовая попытка пробежать километр по вертикали будет интересным испытанием.
После первой попытки мне захотелось продолжить изучение бесконечного, зацикленного ландшафта беговой дорожки, и я провел еще три сеанса ВКонтакте. Я описал каждую из тренировок (также в версии для печати), чтобы вы тоже могли их попробовать. Я рекомендую распечатать копию или держать телефон под рукой на беговой дорожке, чтобы вам не приходилось запоминать точную структуру тренировки. Не забывайте поддерживать хорошую форму: это тяжелые тренировки, поэтому лучше сократить их или отрегулировать наклон в соответствии с вашей текущей физической формой.
Чему я научился на VK Treadmill Challenge?⁃ Во-первых, тренировки на беговой дорожке могут стать хорошей заменой тренировкам в гору, особенно если ваша цель – улучшить технику бега в гору. В моем предыдущем журнальном посте «Техника бега (2): облегчение подъема в гору» обсуждаются некоторые из этих приемов.
⁃ Недавнее исследование показало, что классические переменные на выносливость, такие как VO2max и экономичность бега на склоне +0%, не являются надежными предикторами результативности бега на короткие дистанции (20-30 км), уклон +10% позволяет хорошо охарактеризовать беговые характеристики. Если это так, я готов гнаться за этой эффективностью! Найдите исследование здесь: https://www.ncbi.nlm.nih.gov/pubmed/29.077639
⁃ Разбивка тренировки на этапы или установка протокола интервалов делает ее менее сложной задачей.
⁃ Это самый быстрый и доступный способ пройти настоящий вертикальный километр (ни шагу вниз), если вы не живете в горах.
⁃ Наконец-то я научился понимать, что 15-процентный наклон на беговой дорожке на самом деле имеет большое значение, когда вы выходите на трассу. Час подъема в гору может принести вам захватывающие виды — к сожалению, это не тот случай, когда вы находитесь в тренажерном зале!
Тренировка 1: интервалы подъема в гору VO2MaxВосемь 4-минутных интервалов с уклоном 15%, с 1-минутным отдыхом между каждым интервалом с уклоном 3-5%.
Общая разминка
Строительные упражнения + растяжение (5 минут)Разминка беговой дорожки
10min 1% наклон 10 км/чОсновной набор
Примечание: @hard вы находите сложным, но устойчивым для продолжительности интервала. Темп может меняться между интервалами. RI – интервал восстановления.8 x (4 мин при жестком режиме (уклон 15 % на 7–10 км.
[с более длительным 3-минутным перерывом на восстановление в середине интервалов ]Заминка
3 минуты 1% наклона 10 км/чЗагрузите версию для печати здесь
Тренировка 2: Пирамидальная пороговая сессияИнтервалы восстановления в этой тренировке не будут постепенно увеличиваться, но интервалы восстановления не будут постепенно увеличиваться, но не будут увеличиваться интервалы восстановления в этой тренировке.
Общая разминка
Упражнения на подвижность + растяжка (5 минут)Разминка на беговой дорожке
От 5 до 10 минут с наклоном 1%Основной подход
сложная, но устойчивая на протяжении всего интервала. Темп может меняться между интервалами. RI – интервал восстановления.4 мин 15 % при жестком / 2 мин RI 7 % наклон @ легко
6 мин 15 % @ жестко / 2 мин RI 7 % наклон @ легко
8 мин 15 % @ жестко / 2 мин RI 7% наклона @ easy
10 минут 15 % при жестком / 2 минуты RI 7 % наклона при легком режиме
8 минут 15 % при жестком / 2 минуты RI 7 % наклоне при легком % наклона @легко
4 мин 15% @сложно / 2 мин RI 7% наклона @легкоЗаминка
Тренировка 3: пороговая + силовая сессия
5мин 1% наклона 10км/ч версияЭта тренировка разделена на три этапа, на которых я выполнял три 10-минутных подхода с разным наклоном, заканчивая каждый подход тремя 30-секундными подходами с уклоном 15%.
Общая разминка
Строили мобильности + растяжение (5 минут)Разминка беговой дорожки
5-10-минутного 1%. НаклонОсновной набор (Прогрессивная структура)
I.
10min 8% уклон ~10км.ч (подпорог) + 1 мин. RI (8% 6 км.ч) + 3 x (30с 15% 10-11-12 км.ч / 30с RI 15% 6км.ч) + 2мин RI при 6% 8км.чСтадия II.
10мин 10% наклон ~10км.ч (порог) + 1 мин РИ (8% 6 км.ч) + 3 x (30с 15% 11-12-13 км.ч / 30с РИ 15% 6км.ч) + 2мин РИ при 6% 8км.чЭтап III.
10мин 10% уклон ~10км.ч (порог) + 1 мин РИ (8% 6 км.ч) + 3 x (30с 15% 12-13-14 км.ч / 30с РИ 15% 6км.ч) + 2мин RI при 6% 8 км.чЗаминка
2 мин @легкий темп при 1% 8км.чСкачать версию для печати здесь
Тренировка 4: Threshold + VO2maxsession 9040 Эта тренировка состоит из 1 этапа с тремя более длинными интервалами на первом этапе и 6 более короткими интервалами на втором этапеОбщая разминка
Упражнения на подвижность + растяжка (5 минут)Разминка на беговой дорожке
От 5 до 10 минут с наклоном 1% % уклона 8-10км.
[1 сет: умеренно-тяжелый темп; 2 сет: жесткий темп; 3 набор: очень жесткий темп] – см. анализ сердечного ритмаСтадия II.
6 x (2,5 мин 15 % наклона при пороге + 1 мин 15 % наклона при VO2max + 1,5 мин RI 6 % наклона 7 км.ч)Заминка
2 минуты @легкий темп 1% 10км.чСтатья взята с сайта: www.andriusramonas.com , и исправил Виктору мой лучший вид «Продолжай».
«…так что вы ничего не делаете, кроме диеты и упражнений как сумасшедшие; у вас нет жизни, но вы сбрасываете 160 фунтов. Или вы не делаете ничего радикального, наслаждаетесь собой и в конечном итоге весите 170 фунтов» , немного выше вашей цели. Что бы вы предпочли, Джонатан?»
Ага. Поэтому BAT создает симметричные биполярные источники питания, которые позволяют их оборудованию дышать. (Симметричный означает, что источник питания генерирует как положительные, так и отрицательные шины 150 В, а не полагается на землю — полную schmutz , как это часто бывает — для возврата.
Виктор: «Мы также считаем, что триодные схемы с пластинчатой нагрузкой всегда звучат лучше, поэтому на выходе VK-P10 нет катодных повторителей. И нет каскодных схем. Тем не менее, выходное сопротивление фонокорректора достаточно низкое, чтобы быть проблемой в «обычных» системах. Под этим я подразумеваю типичный предусилитель линейного каскада, такой как наш VK-5, с входным сопротивлением не менее 10 кОм и межсоединением менее 100 футов или около того».
Рекомендации по сборке
Я был удивлен, узнав, что VK-P10 изготовлен из оцинкованной стали, обладающей магнитными свойствами. По словам Виктора, это обеспечивает некоторую степень ослабления составляющей 60 Гц, которая окружает всех нас. (Нью-Йорк — такая излучаемая среда, удивительно, что мы иногда не просыпаемся утром с тостами. Возможно, так и есть…)Я узнал, что эта чувствительность сборки отражена повсюду в дизайне. Например, массовое демпфирование верхней крышки шасси может придать устройству более существенное «ощущение», но не имеет большого реального эффекта.
BAT также использует керамические патроны с серебряными контактами, и… этот список можно продолжать и продолжать. Я настоятельно рекомендую всем заинтересованным сторонам обращаться в BAT за копиями их различных официальных документов. Они проделывают потрясающую работу, очень убедительно объясняя инженерные решения и реализации, представленные в их проектах. Это действительно увлекательное чтение.
Универсальность
Входной каскад включает в себя большинство элементов, обеспечивающих универсальность VK-P10. Это включает выбираемую пользователем загрузку картриджа сопротивлением (100, 1 кОм или 10 кОм или по выбору пользователя) и емкостью (100 пикофарад, 470 пФ, 1000 пФ или по выбору пользователя) или любую параллельную комбинацию.Имеется переключатель для выбора между режимами высокого и низкого усиления. Low снижает выходной сигнал на 6 дБ для картриджей с форсажной камерой. Есть еще один переключатель для выбора между режимами Direct и Step-Up, который вводит пару трансформаторов в выходы. В трансиверах есть отводы на 12 и 18 дБ, которые выбираются… конечно, нажатием переключателя!
В целом, это дает пользователю диапазон усиления от 50 до 83 дБ для работы. Фонокорректор должен работать с картриджами мощностью от 0,1 мВ до 5 мВ. Если вы не можете заставить свой картридж прижиться, откажитесь от него.
Загрузка
Поскольку VK-P10 такой гибкий, я возился с его многочисленными настройками. Я попробовал установленные на плате значения 100, 10 кОм и 100 кОм.Садитесь на Симфоническую линию RG-8. Ну… пока оставим это здесь. Он любит небольшую нагрузку, но не очень доволен на 100 Ом. Я предпочел настройку 1 кОм, которую использовал на протяжении всего периода испытаний, без включения емкости. Входное сопротивление 10 кОм или 47 кОм без нагрузки было даже лучше, но слишком шумно для Нью-Йорка с высоким уровнем радиопомех. Лучше оставить все как есть, что?
Звук
Я посмотрел на систему, когда сел, чтобы критически прослушать.Большую часть периода обзора я остановился на RG-8. Он отлично сочетался с VK-P10. Картридж звучал впечатляюще при начальной массе 1,4 г VTF. С мазком Mortite на шелле осязаемость и «тело» улучшились. (Для этой цели я держу под рукой мортитовые козявки весом 1/10 грамма. Можете ли вы сказать: «Получи жизнь!»?)
Подклассы и аллотипы IgG: от структуры к эффекторным функциям
1. Schur PH. подклассы IgG. Историческая перспектива. Monogr Allergy (1988) 23:1–11 [PubMed] [Google Scholar]
2. Bruhns P, Iannascoli B, England P, Mancardi DA, Fernandez N, Jorieux S, et al. Специфичность и аффинность рецепторов Fcgamma человека и их полиморфных вариантов для подклассов IgG человека. Кровь (2009) 113 (16): 3716–25.
3. Pan Q, Hammarstrom L. Молекулярная основа дефицита подкласса IgG. Immunol Rev (2000) 178:99–110. 10.1034/j.1600-065X.2000.17815.x [PubMed] [CrossRef] [Google Scholar]
4. Влуг А., Ньювенхейс Э.Дж., ван Эйк Р.В., Герцен Х.Г., ван Хаут А.Дж. Нефелометрические измерения подклассов IgG человека и их референтные диапазоны. Энн Биол Клин (Париж) (1994) 52 (7–8): 561–7. [PubMed] [Google Scholar]
5. Pone EJ, Zhang J, Mai T, White CA, Li G, Sakakura JK, et al. Передача сигналов BCR синергизирует с передачей сигналов TLR для индукции AID и переключения класса иммуноглобулинов через неканонический путь NF-kappaB. Нат Коммун (2012) 3:767. 10.1038/ncomms1769[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Pone EJ, Zan H, Zhang J, Al-Qahtani A, Xu Z, Casali P. Toll-подобные рецепторы и рецепторы B-клеток взаимодействуют для индуцировать рекомбинацию ДНК с переключением класса иммуноглобулинов: отношение к реакциям микробных антител.
7. Berkowska MA, Driessen GJ, Bikos V, Grosserichter-Wagener C, Stamatopoulos K, Cerutti A, et al. . В-клетки памяти человека происходят из трех различных путей созревания, зависимых от зародышевого центра и независимых от них. Кровь (2011) 118 (8): 2150–8. 10.1182/кровь-2011-04-345579[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Aalberse RC, Stapel SO, Schuurman J, Rispens T. Иммуноглобулин G4: нечетное антитело. Clin Exp Allergy (2009) 39(4):469–77. 10.1111/j.1365-2222.2009.03207.x [PubMed] [CrossRef] [Google Scholar]
9. Ferrante A, Beard LJ, Feldman RG. Распределение подклассов IgG антител к бактериальным и вирусным антигенам. Pediatr Infect Dis J (1990) 9 (8 Suppl): S16–24. 10.1097/00006454-1901-00004 [PubMed] [CrossRef] [Google Scholar]
10. Джефферис Р., Кумараратне Д.С. Селективный дефицит подкласса IgG: количественная оценка и клиническая значимость.
11. Siber GR, Schur PH, Aisenberg AC, Weitzman SA, Schiffman G. Корреляция между IgG- 2 концентрации и ответ антител на бактериальные полисахаридные антигены. N Engl J Med (1980) 303 (4): 178–82. 10.1056/NEJM198007243030402 [PubMed] [CrossRef] [Google Scholar]
12. Барретт Д.Дж., Аюб Э.М. Ограничение антител подкласса IgG2 к пневмококковым полисахаридам. Clin Exp Immunol (1986) 63(1):127–34. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Schauer U, Stemberg F, Rieger CH, Buttner W, Borte M, Schubert S, et al. Уровни антител, специфичных к столбнячному анатоксину, Haemophilus influenzae типа b и пневмококковому капсульному полисахариду у здоровых детей и взрослых. Clin Diagn Lab Immunol (2003) 10(2):202–7. 10.1128/CDLI.10.2.202-207.2003 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
14. Хаммарстром Л.
15. Hammarstrom L, Carbonara AO, DeMarchi M, Lefranc G, Moller G, Smith CI, et al. Характер рестрикции подкласса антиген-специфических антител у доноров с дефектной экспрессией генов константной области тяжелой цепи подкласса IgG или IgA. Клин Иммунол Иммунопатол (1987) 45(3):461–70. 10.1016/0090-1229(87)-3 [PubMed] [CrossRef] [Google Scholar]
16. Kuijpers TW, Weening RS, Out TA. Дефицит подкласса IgG и рецидивирующие пиогенные инфекции, невосприимчивость к бактериальным полисахаридным антигенам. Allergol Immunopathol (Madr) (1992) 20(1):28–34. [PubMed] [Google Scholar]
17. Латифф А.Х., Керр М.А. Клиническое значение дефицита иммуноглобулина А. Энн Клин Биохим (2007) 44 (Pt 2): 131–9. 10.1258/000456307780117993 [PubMed] [CrossRef] [Google Scholar]
18.
19. Vidarsson G, Sigurdardottir ST, Gudnason T, Kjartansson S, Kristinsson KG, Ingolfsdottir G, et al. Изотипы и опсонофагоцитоз антител против пневмококка типа 6В, вызванных у младенцев и взрослых экспериментальной вакциной против пневмококка типа 6В и столбнячным анатоксином. Заразить Иммуна (1998) 66(6):2866–70. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Stapleton NM, Andersen JT, Stemerding AM, Bjarnarson SP, Verheul RC, Gerritsen J, et al. Конкуренция за FcRn-опосредованный транспорт приводит к короткому периоду полужизни человеческого IgG3 и предлагает терапевтический потенциал. Нат Коммун (2011) 2:599. 10.1038/ncomms1608 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21.
22. Хассан М.С., Ислам К.Б., Хаммарстром Л., Смит С.И. Регуляция экспрессии C гамма 3. Роль переключения в связанных с аллотипом вариациях уровней IgG3 в сыворотке человека. J Immunol (1992) 148(8):2555–62. [PubMed] [Google Scholar]
23. Brezski RJ, Jordan RE. Расщепление IgG протеазами, связанными с инвазивными заболеваниями: тактика уклонения от иммунитета хозяина? MAbs (2010) 2(3):212–20. 10.4161/mabs.2.3.11780 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Soderstrom T, Enskog A, Samuelsson BE, Cedergren B. Иммуноглобулиновый подкласс (IgG3) ограничение анти-P и анти-Pk антител у пациентов с редкой p группой крови. J Immunol (1985) 134(1):1–3. [PubMed] [Google Scholar]
25. Brouwers HA, Overbeeke MA, Ouwehand WH, Keuning K, van Ertbruggen I, van Leeuwen EF, et al.
26. Mawas F, Wiener E, Williamson LM, Rodeck CH. Подклассы иммуноглобулина G антигена тромбоцитов человека 1a в материнской сыворотке: связь с тяжестью неонатальной аллоиммунной тромбоцитопении. Eur J Haematol (1997) 59(5):287–92. 10.1111/j.1600-0609.1997.tb01688.x [PubMed] [CrossRef] [Google Scholar]
27. Поллок Дж.М., Боумен Дж.М. Подклассы анти-Rh(D)IgG и тяжесть резус-гемолитической болезни новорожденных. Вокс Санг (1990) 59(3):176–9. 10.1111/j.1423-0410.1990.tb00854.x [PubMed] [CrossRef] [Google Scholar]
28. Мейц И., Боссайт X, Проесманс М., Бек К.Д. Изолированный дефицит IgG3 у детей: лечить или не лечить? Презентация случая и обзор литературы.
29. Aalberse RC, van der Gaag R, van Leeuwen J. Серологические аспекты антител IgG4. I. Длительная иммунизация приводит к ограниченному IgG4 ответу. Джей Иммунол (1983) 130:722–6. [PubMed] [Google Scholar]
30. Aalberse RC, Dieges PH, Knul-Bretlova V, Vooren P, Aalbers M, van Leeuwen J. IgG4 в качестве блокирующего антитела. Clin Rev Allergy (1983) 1(2):289–302 [PubMed] [Google Scholar]
31. Nouri-Aria KT, Wachholz PA, Francis JN, Jacobson MR, Walker SM, Wilcock LK, et al. Иммунотерапия пыльцой трав индуцирует слизистые и периферические реакции IL-10 и блокирует активность IgG. J Immunol (2004) 172(5):3252–9. 10.4049/jimmunol.172.5.3252 [PubMed] [CrossRef] [Google Scholar]
32. Ютель М., Акдис СА. Иммунологические механизмы аллергенспецифической иммунотерапии. Аллергия (2011) 66(6):725–32. 10.1111/j.1398-9995.2011.02589.x [PubMed] [CrossRef] [Google Scholar]
33.
34. Андерсен Б.Р., Терри В.Д. Гамма-G4-глобулиновое антитело, вызывающее ингибирование фактора свертывания крови VIII. Природа (1968) 217 (5124): 174–5. 10.1038/217174a0 [CrossRef] [Google Scholar]
35. Iizuka A, Nagao T. Анализ подклассов тяжелых цепей IgG аллоантител к фактору IX с помощью перекрестного иммуноэлектрофореза фактора IX с использованием метода промежуточного геля. Br J Haematol (1983) 53(4):687–8. 10.1111/j.1365-2141.1983.tb07323.x [PubMed] [CrossRef] [Google Scholar]
36. van Schouwenburg PA, Kriekaert CL, Nurmohamed M, Hart M, Rispens T, Aarden L, et al. Продукция IgG4 против адалимумаба при длительном лечении больных РА. J Clin Immunol (2012) 32:1000–6.
37. Ottesen EA, Skvaril F, Tripathy SP, Poindexter RW, Hussain R. Известность IgG4 в ответе антител IgG на человеческий филяриатоз. J Immunol (1985) 134(4):2707–12. [PubMed] [Google Scholar]
38. Adjobimey T, Hoerauf A. Индукция иммуноглобулина G4 при филяриатозе человека: показатель иммунорегуляции. Ann Trop Med Parasitol (2010) 104(6):455–64. 10.1179/136485910X127863898
[бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Kurniawan A, Yazdanbakhsh M, van Ree R, Aalberse R, Selkirk ME, Partono F, et al. Дифференциальная экспрессия специфических ответов антител IgE и IgG4 при бессимптомном и хроническом филяриатозе человека. J Immunol (1993) 150(9):3941–50. [PubMed] [Google Scholar]
40. Sah RP, Chari ST. Серологические проблемы при системном заболевании, связанном с IgG4, и аутоиммунном панкреатите. Curr Opin Rheumatol (2011) 23(1):108–13. 10.1097/BOR.0b013e3283413469 [PubMed] [CrossRef] [Google Scholar]
41.
42. Махаджан В.С., Матту Х., Дешпанде В., Пиллаи С.С., Стоун Д.Х. Заболевание, связанное с IgG4. Анну Рев Патол (2014) 9: 315–47. 10.1146/annurev-pathol-012513-104708 [PubMed] [CrossRef] [Google Scholar]
43. Поттер М. Структурные корреляты разнообразия иммуноглобулинов. Surv Immunol Res (1983) 2(1):27–42 [PubMed] [Google Scholar]
44. Wu TT, Johnson G, Kabat EA. Распределение длины CDRh4 в антителах. Белки (1993) 16(1):1–7. 10.1002/prot.340160102 [PubMed] [CrossRef] [Google Scholar]
45. Kabat EA, Wu TT, Bilofsky H. Гены вариабельной области каркаса иммуноглобулина собираются из небольших сегментов ДНК – гипотеза. Proc Natl Acad Sci USA (1978) 75(5):2429–33.
46. Ferrara C, Grau S, Jager C, Sondermann P, Brunker P, Waldhauer I, et al. Уникальные углевод-углеводные взаимодействия необходимы для высокоаффинного связывания между Fc{gamma}RIII и антителами, лишенными сердцевинной фукозы. Proc Natl Acad Sci USA (2011) 108(31):12669–74. 10.1073/pnas.1108455108 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47. Радаев С., Мотыка С., Фридман В.Х., Сотес-Фридман С., Сун П.Д. Структура рецептора Fcgamma типа III человека в комплексе с Fc. J Biol Chem (2001) 276(19):16469–77. 10.1074/jbc.M100350200 [PubMed] [CrossRef] [Google Scholar]
48. Kato K, Sautes-Fridman C, Yamada W, Kobayashi K, Uchiyama S, Kim H, et al. Структурные основы взаимодействия рецепторов IgG и Fcgamma. Дж Мол Биол (2000) 295(2):213–24. 10.1006/jmbi.1999.3351 [PubMed] [CrossRef] [Google Scholar]
49. Радаев С., Сун П. Распознавание иммуноглобулинов рецепторами Fcgamma.
50. Sondermann P, Pincetic A, Maamary J, Lammens K, Ravetch JV. Общий механизм модуляции эффекторной функции иммуноглобулина. Proc Natl Acad Sci U S A (2013) 110(24):9868–72. 10.1073/pnas.1307864110 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51. Уэст А.П., младший, Бьоркман П.Дж. Кристаллическая структура и свойства связывания иммуноглобулина G рецептора Fc, связанного с главным комплексом гистосовместимости человека (,). Биохимия (2000) 39(32):9698–708. 10.1021/bi000749m [PubMed] [CrossRef] [Google Scholar]
52. Burmeister WP, Huber AH, Bjorkman PJ. Кристаллическая структура комплекса неонатального Fc-рецептора крысы с Fc. Природа (1994) 372 (6504): 379–83. 10.1038/372379a0 [PubMed] [CrossRef] [Google Scholar]
53. Кибл А.Х., Хан З., Форстер А., Джеймс Л.С. TRIM21 представляет собой рецептор IgG, который структурно, термодинамически и кинетически консервативен.
54. Kapur R, Kustiawan I, Vestrheim A, Koeleman C, Visser R, Einarsdottir HK, et al. Заметное отсутствие фукозилирования IgG1-Fc аллоантител тромбоцитов при беременности. Кровь (2014) 123 (4): 471–80. 10.1182/blood-2013-09-527978 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
55. Wuhrer M, Porcelijn L, Kapur R, Koeleman CA, Deelder A, de Haas M, et al. Регулируемые паттерны гликозилирования IgG во время аллоиммунных ответов против антигенов тромбоцитов человека. Джей Протеом Рез (2009 г.)) 8(2):450–6. 10.1021/pr800651j [PubMed] [CrossRef] [Google Scholar]
56. Райт А., Тао М.Х., Кабат Э.А., Моррисон С.Л. Гликозилирование вариабельной области антител: влияние положения на связывание антигена и структуру углеводов. EMBO J (1991) 10 (10): 2717–23. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Ackerman ME, Crispin M, Yu X, Baruah K, Boesch AW, Harvey DJ, et al.
58. Guhr T, Bloem J, Derksen NI, Wuhrer M, Koenderman AH, Aalberse RC, et al. Обогащение сиалилированного IgG путем лектинового фракционирования не повышает эффективность иммуноглобулина G в мышиной модели иммунной тромбоцитопении. PLoS One (2011) 6(6):e21246. 10.1371/journal.pone.0021246 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
59. Stadlmann J, Weber A, Pabst M, Anderle H, Kunert R, Ehrlich HJ, et al. Внимательный взгляд на сиалирование человеческого IgG и распределение подклассов после фракционирования лектина. Протеомика (2009 г.) 9(17):4143–53. 10.1002/pmic.200800931 [PubMed] [CrossRef] [Google Scholar]
60. Radcliffe CM, Arnold JN, Suter DM, Wormald MR, Harvey DJ, Royle L, et al. Клетки фолликулярной лимфомы человека содержат олигоманнозные гликаны в антигенсвязывающем участке В-клеточного рецептора.
[PubMed] [CrossRef] [Google Scholar]
61. Hamilton RG. Измерения подкласса человеческого IgG в клинической лаборатории. Clin Chem (1987) 33 (10): 1707–25. [PubMed] [Академия Google]
62. Ру К.Х., Стрелец Л., Михаэльсен Т.Е. Гибкость подклассов IgG человека. J Immunol (1997) 159(7):3372–82. [PubMed] [Google Scholar]
63. Michaelsen TE, Naess LM, Aase A. Человеческий IgG3 снижен, а IgG1, IgG2 и IgG4 не изменились по размеру молекулы за счет мягкого восстановления и повторного окисления без каких-либо серьезных изменений в эффекторных функциях. Мол Иммунол (1993) 30(1):35–45. 10.1016/0161-5890(93)-A [PubMed] [CrossRef] [Google Scholar]
64. Saluk PH, Clem LW. Уникальная молекулярная масса тяжелой цепи IgG3 человека. Джей Иммунол (1971) 107(1):298–301 [PubMed] [Google Scholar]
65. Carrasco B, Garcia de la Torre J, Davis KG, Jones S, Athwal D, Walters C, et al. Кристаллогидродинамика для решения проблемы гидратации многодоменных белков: открытые физиологические конформации IgG человека.
66. Wypych J, Li M, Guo A, Zhang Z, Martinez T, Allen MJ, et al. Человеческие антитела IgG2 демонстрируют структурные изоформы, опосредованные дисульфидом. Дж. Биол. Химия (2008) 283 (23): 16194–205. 10.1074/jbc.M709987200 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
67. Zhang B, Harder AG, Connelly HM, Maheu LL, Cockrill SL. Определение дисульфидной связности Fab-шарнира в структурных изоформах рекомбинантного человеческого антитела иммуноглобулина G2. Анальная химия (2010) 82 (3): 1090–9. 10.1021/ac6z [PubMed] [CrossRef] [Google Scholar]
68. Dillon TM, Ricci MS, Vezina C, Flynn GC, Liu YD, Rehder DS, et al. Структурно-функциональная характеристика изоформ дисульфидов подкласса IgG2 человека. J Biol Chem (2008) 283(23):16206–15. 10.1074/jbc.M709988200 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
69. Lightle S, Aykent S, Lacher N, Mitaksov V, Wells K, Zobel J, et al.
70. Yoo EM, Wims LA, Chan LA, Morrison SL. Человеческий IgG2 может образовывать ковалентные димеры. J Immunol (2003) 170(6):3134–8. 10.4049/jimmunol.170.6.3134 [PubMed] [CrossRef] [Google Scholar]
71. Chivers PT, Prehoda KE, Raines RT. Мотив CXXC: реостат в активном центре. Биохимия (1997) 36(14):4061–6. 10.1021/bi9628580 [PubMed] [CrossRef] [Google Scholar]
72. Bloom JW, Madanat MS, Marriott D, Wong T, Chan SY. Внутрицепочечная дисульфидная связь в центральной шарнирной области IgG4 человека. Protein Sci (1997) 6(2):407–15. 10.1002/pro.5560060217 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
73. Angal S, King DJ, Bodmer MW, Turner A, Lawson AD, Roberts G, et al. Замена одной аминокислоты устраняет гетерогенность химерного антитела мыши/человека (IgG4).
74. Rispens T, Ooijevaar-de Heer P, Bende O, Aalberse RC. Механизм обмена Fab-плеч иммуноглобулина G4. J Am Chem Soc (2011) 133 (26): 10302–11. 10.1021/ja203638y [PubMed] [CrossRef] [Google Scholar]
75. Van der Zee JS, Van Swieten P, Aalberse RC. Серологические аспекты антител IgG4. II. Антитела IgG4 образуют небольшие непреципитирующие иммунные комплексы благодаря функциональной моновалентности. Джей Иммунол (1986) 137(11):3566–71. [PubMed] [Google Scholar]
76. van der Neut KM, Schuurman J, Losen M, Bleeker WK, Martinez-Martinez P, Vermeulen E, et al. Противовоспалительная активность антител IgG4 человека при динамическом обмене плечами Fab. Наука (2007) 317 (5844): 1554–7. 10.1126/science.1144603 [PubMed] [CrossRef] [Google Scholar]
77. Rispens T, Davies AM, Ooijevaar-de Heer P, Absalah S, Bende O, Sutton BJ, et al. Динамика взаимодействий между тяжелыми цепями в подклассах человеческого иммуноглобулина G (IgG), изученная с помощью кинетического обмена плечами Fab.
78. Labrijn AF, Rispens T, Meesters J, Rose RJ, den Bleker TH, Loverix S, et al. Видоспецифичные детерминанты в домене IgG Ch4 обеспечивают обмен Fab-плечами, влияя на силу нековалентного взаимодействия Ch4-Ch4. J Immunol (2011) 187(6):3238–46. 10.4049/jimmunol.1003336 [PubMed] [CrossRef] [Google Scholar]
79. Labrijn AF, Buijsse AO, van den Bremer ET, Verwilligen AY, Bleeker WK, Thorpe SJ, et al. Терапевтические антитела IgG4 участвуют в обмене Fab-плеч с эндогенным человеческим IgG4 in vivo. Нат Биотехнолог (2009 г.)) 27(8):767–71. 10.1038/nbt.1553 [PubMed] [CrossRef] [Google Scholar]
80. Lefranc MP, Lefranc G. Аллотипы Gm, Km и Am человека и их молекулярная характеристика: замечательная демонстрация полиморфизма. Методы Мол Биол (2012) 882:635–80. 10.1007/978-1-61779-842-9_34 [PubMed] [CrossRef] [Google Scholar]
81. де Ланге Г.
82. Джефферис Р., Лефранк, член парламента. Аллотипы иммуноглобулинов человека: возможные последствия для иммуногенности. MAbs (2009 г.)) 1(4):332-8. 10.4161/mabs.1.4.9122 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
83. Dard P, Lefranc MP, Osipova L, Sanchez-Mazas A. Изменчивость последовательности ДНК аллелей IGHG3, связанных с основным Гаплотипы G3m в популяциях человека. Eur J Hum Genet (2001) 9 (10): 765–72. 10.1038/sj.ejhg.5200700 [PubMed] [CrossRef] [Google Scholar]
84. Brusco A, Saviozzi S, Cinque F, DeMarchi M, Boccazzi C, De LG, et al. Молекулярная характеристика изоаллотипов гена иммуноглобулина G4. Евр Дж Иммуногенет (1998) 25(5):349–55. 10.1111/j.1744-313X.1998.tb01152.x [PubMed] [CrossRef] [Google Scholar]
85. Brusco A, de Lange GG, Boccazzi C, Carbonara AO. Молекулярная характеристика аллотипов Gm (n+) и G2m (n-).
86. de Lange G. Моноклональные антитела против аллотипов иммуноглобулинов человека. Кандидат наук. диссертация, Лондонский университет, Лондон (1988). [Google Scholar]
87. Fudenberg HH, Fudenberg BR. Антитело к наследственному человеческому фактору гамма-глобулина (Gm), возникающему в результате несовместимости матери и плода. Наука (1964) 145(3628):170–1. 10.1126/science.145.3628.170 [PubMed] [CrossRef] [Google Scholar]
88. de Lange G, Zhong FM, Henke J, Feng ZC, Bernhardt R, van Leeuwen F, et al. Аллотипы иммуноглобулинов в китайской популяции: сравнение частот гаплотипов с другими азиатскими группами. J Immunogenet (1985) 12(4–5):191–5. 10.1111/j.1744-313X.1985.tb00846.x [PubMed] [CrossRef] [Google Scholar]
89. Goetze AM, Zhang Z, Liu L, Jacobsen FW, Flynn GC. Быстрый скрининг LC-MS на наличие модификаций Fc IgG и аллельных вариантов в крови. Мол Иммунол (2011) 49(1–2): 338–52.
90. Cook CE, Steinberg AG. Аминокислотная замена в цепи гамма-1 иммуноглобулина G человека, ассоциированная с аллотипом Gm(2). Мол Иммунол (1979) 16(8):555–8. 10.1016/0161-5890(79)
91. Heilig R, Eckenberg R, Petit JL, Fonknechten N, Da Silva C, Cattolico L, et al. Последовательность ДНК и анализ хромосомы 14 человека. Nature (2003) 421 (6923): 601–7. 10.1038/nature01348 [PubMed] [CrossRef] [Google Scholar]
92. Hougs L, Garred P, Kawasaki T, Kawasaki N, Svejgaard A, Barington T. Три новых аллеля IGHG2 и их распространенность у датских европеоидов, негров Мозамбика и японцев. Тканевые антигены (2003) 61(3):231–9. 10.1034/j.1399-0039.2003.00048.x [PubMed] [CrossRef] [Google Scholar]
93. Bartelds GM, de Groot E, Nurmohamed MT, Hart MH, van Eede PH, Wijbrandts CA, et al. Неожиданная отрицательная связь между несоответствием аллотипов IgG1 и образованием антиадалимумаба: когортное исследование 1.
94. Magdelaine-Beuzelin C, Vermeire S, Goodall M, Baert F, Noman M, Assche GV, et al. Полиморфизм гена, кодирующего тяжелую цепь IgG1 (аллотипы G1m), и развитие антител к инфликсимабу 8. Pharmacogenet Genomics (2009) 19(5):383–7. 10.1097/FPC.0b013e32832a06bf [PubMed] [CrossRef] [Google Scholar]
95. Морелл А., Скварил Ф., Стейнберг А.Г., Ван Логем Э., Терри В.Д. Корреляции между концентрациями четырех подклассов аллотипов IgG и Gm в сыворотке крови человека. Джей Иммунол (1972) 108(1):195–206 [PubMed] [Google Scholar]
96. Iserentant D, Fiers W. Вторичная структура мРНК и эффективность инициации трансляции. Джин (1980) 9 (1–2): 1–12. 10.1016/0378-1119(80)
-8 [PubMed] [CrossRef] [Google Scholar]97. Mao Y, Liu H, Liu Y, Tao S. Расшифровка правил, по которым динамика вторичной структуры мРНК влияет на эффективность трансляции у Saccharomyces cerevisiae.
98. Pan Q, Petit-Frere C, Hammarstrom L. Ассоциированный с аллотипом полиморфизм в промоторе gamma3 определяет скорость транскрипции gamma3 зародышевой линии, но не влияет на переключение и последующую продукцию IgG3. Eur J Immunol (2000) 30(8):2388–93. 10.1002/1521-4141(2000)30:8<2388::AID-IMMU2388>3.0.CO;2-C [PubMed] [CrossRef] [Google Scholar]
99. Diebolder C, Beurskens FJ, de Jong RN, Конинг Р.И., Струман К., Линдорфер М. и соавт. Комплемент активируется гексамерами IgG, собранными на поверхности клетки. Наука (2014) 343 (6176): 1260–3. 10.1126/наука.1248943 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
100. Zauner G, Selman MH, Bondt A, Rombouts Y, Blank D, Deelder AM, et al. Гликопротеомный анализ антител. Молекулярная клеточная протеомика (2013) 12(4):856–65. 10.1074/mcp.R112.026005 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
101.
102. Bondt A, Rombouts Y, Selman MH, Hensbergen PJ, Reiding KR, Hazes JM, et al. Анализ гликозилирования IgG Fab с использованием нового масс-спектрометрического высокопроизводительного метода профилирования выявляет изменения, связанные с беременностью. Мол клеточная протеомика (2014). 10.1074/mcp.M114.039537 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
103. Kapur R, Della Valle L, Sonneveld M, Hipgrave Ederveen A, Visser R, Ligthart P, et al. Низкое анти-RhD IgG-Fc-фукозилирование при беременности: новая переменная, предсказывающая тяжесть гемолитической болезни плода и новорожденного. Бр Дж. Гематол (2014) 166 (6): 936–45. 10.1111/bjh.12965 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
104.
105. Chen G, Wang Y, Qiu L, Qin X, Liu H, Wang X, et al. Профилирование Fc-гликозилирования IgG человека выявляет ассоциации с возрастом, полом, женскими половыми гормонами и раком щитовидной железы. J Протеомика (2012) 75 (10): 2824–34. 10.1016/j.jprot.2012.02.001 [PubMed] [CrossRef] [Google Scholar]
106. Wang J, Balog CI, Stavenhagen K, Koeleman CA, Scherer HU, Selman MH, et al. Fc-гликозилирование IgG1 модулируется стимулами В-клеток. Mol Cell Proteomics (2011) 10(5):M110. 10.1074/mcp.M110.004655 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
107. Prados MB, La Blunda J, Szekeres-Bartho J, Caramelo J, Miranda S. Прогестерон вызывает переключение олигосахарилтрансферазы экспрессия изоформы: влияние на N-гликозилирование IgG.
108. Bowden TA, Baruah K, Coles CH, Harvey DJ, Yu X, Song BD, et al. Химический и структурный анализ промежуточного соединения сворачивания антител, захваченного во время биосинтеза гликанов. J Am Chem Soc (2012) 134 (42): 17554–63. 10.1021/ja306068g [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
109. Mimura Y, Sondermann P, Ghirlando R, Lund J, Young SP, Goodall M, et al. Роль олигосахаридных остатков IgG1-Fc в связывании Fc гамма RIIb. J Biol Chem (2001) 276(49):45539–47. 10.1074/jbc.M107478200 [PubMed] [CrossRef] [Google Scholar]
110. Sondermann P, Huber R, Oosthuizen V, Jacob U. Кристаллическая структура 3.2-A комплекса фрагмент Fc IgG1 человека-Fc gammaRIII. Природа (2000) 406 (6793): 267–73. 10.1038/35018508 [PubMed] [CrossRef] [Google Scholar]
111. Mizushima T, Yagi H, Takemoto E, Shibata-Koyama M, Isoda Y, Iida S, et al. Структурная основа повышения эффективности терапевтических антител при дефукозилировании их Fc-гликанов.
112. Shields RL, Lai J, Keck R, O’Connell LY, Hong K, Meng YG, et al. Отсутствие фукозы в N-связанном олигосахариде IgG1 человека улучшает связывание с человеческим Fcgamma RIII и антителозависимую клеточную токсичность. J Biol Chem (2002) 277(30):26733–40. 10.1074/jbc.M202069200 [PubMed] [CrossRef] [Google Scholar]
113. Yamane-Ohnuki N, Satoh M. Производство терапевтических антител с контролируемым фукозилированием. MAbs (2009) 1(3):230–6. 10.4161/mabs.1.3.8328 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
114. Джефферис Р. Терапия рекомбинантными антителами: влияние гликозилирования на механизмы действия. Trends Pharmacol Sci (2009) 30(7):356–62. 10.1016/j.tips.2009.04.007 [PubMed] [CrossRef] [Google Scholar]
115. Shibata-Koyama M, Iida S, Misaka H, Mori K, Yano K, Shitara K, et al. Нефукозилированный ритуксимаб потенцирует фагоцитоз нейтрофилов человека за счет высокой степени связывания с экспрессией FcgammaRIIIb и МНС класса II на фагоцитирующих нейтрофилах.
116. Peipp M, Lammerts van Bueren JJ, Schneider-Merck T, Bleeker WK, Dechant M, Beyer T, et al. Фукозилирование антител по-разному влияет на цитотоксичность, опосредованную эффекторными клетками NK и PMN. Кровь (2008) 112 (6): 2390–9. 10.1182/blood-2008-03-144600 [PubMed] [CrossRef] [Google Scholar]
117. Selman MH, de Jong SE, Soonawala D, Kroon FP, Adegnika AA, Deelder AM, et al. Изменения антигенспецифического N-гликозилирования Fc IgG1 при вакцинации против гриппа и столбняка. Mol Cell Proteomics (2012) 11(4):M111. 10.1074/mcp.M111.014563 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
118. Кодар К., Штадльманн Дж., Клаамас К., Сергеев Б., Куртенков О. Профилирование N-гликанов Fc иммуноглобулина G у пациентов с раком желудка с помощью LC-ESI-MS: связь с прогрессированием опухоли и выживаемостью. Glycoconj J (2012) 29(1):57–66. 10.1007/s10719-011-9364-z [PubMed] [CrossRef] [Google Scholar]
119.
120. Bakovic MP, Selman MH, Hoffmann M, Rudan I, Campbell H, Deelder AM, et al. Высокопроизводительное профилирование N-гликозилирования IgG Fc с помощью масс-спектрометрии гликопептидов. J Proteome Res (2013) 12(2):821–31. 10.1021/pr300887z [PubMed] [CrossRef] [Google Scholar]
121. Ferrara C, Brunker P, Suter T, Moser S, Puntener U, Umana P. Модуляция эффекторных функций терапевтических антител с помощью инженерии гликозилирования: влияние локализации фермента Гольджи домен и коэкспрессия гетерологичной бета1, 4-N-ацетилглюкозаминилтрансферазы III и альфа-маннозидазы Гольджи II. Биотехнология Биоэнг (2006) 93(5):851–61.
122. Капур Р., Эйнарсдоттир Х.К., Видарссон Г. IgG-эффекторные функции: «хорошее, плохое и уродливое». Immunol Lett (2014) 160(2):139–44. 10.1016/j.imlet.2014.01.015 [PubMed] [CrossRef] [Google Scholar]
123. Bondt A, Selman MH, Deelder AM, Hazes JM, Willemsen SP, Wuhrer M, et al. Связь между галактозилированием иммуноглобулина G и улучшением течения ревматоидного артрита во время беременности не зависит от сиалирования. J Proteome Res (2013) 12(10):4522–31. 10.1021/пр400589m [PubMed] [CrossRef] [Google Scholar]
124. Ahmed AA, Giddens J, Pincetic A, Lomino JV, Ravetch JV, Wang LX, et al. Структурная характеристика противовоспалительных иммуноглобулинов fc белков. J Mol Biol (2014) 426 (18): 3166–79. 10.1016/j.jmb.2014.07.006 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
125. Kaneko Y, Nimmerjahn F, Ravetch JV. Противовоспалительная активность иммуноглобулина G в результате сиалирования Fc.
126. Anthony RM, Nimmerjahn F, Ashline DJ, Reinhold VN, Paulson JC, Ravetch JV. Резюме противовоспалительной активности IVIG с рекомбинантным Fc IgG. Наука (2008) 320 (5874): 373–6. 10.1126/science.1154315 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
127. Sarma JV, Ward PA. Система комплемента. Cell Tissue Res (2011) 343 (1): 227–35. 10.1007/s00441-010-1034-0 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
128. Bindon CI, Hale G, Bruggemann M, Waldmann H. Изотипы моноклональных IgG человека различаются по функции активации комплемента на уровне C4, а также C1q. J Exp Med (1988) 168 (1): 127–42. 10.1084/jem.168.1.127 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
129. Schumaker VN, Calcott MA, Spiegelberg HL, Muller-Eberhard HJ. Ультрацентрифугные исследования связывания IgG разных подклассов с субъединицей Clq первого компонента комплемента.
130. Тао М.Х., Смит Р.И., Моррисон С.Л. Структурные особенности иммуноглобулина G человека, определяющие изотип-специфические различия в активации комплемента. J Exp Med (1993) 178(2):661–7. 10.1084/jem.178.2.661 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
131. Idusogie EE, Presta LG, Gazzano-Santoro H, Totpal K, Wong PY, Ultsch M, et al. Картирование сайта связывания C1q на ритуксане, химерном антителе с Fc IgG1 человека. J Immunol (2000) 164(8):4178–84. 10.4049/jimmunol.164.8.4178 [PubMed] [CrossRef] [Google Scholar]
132. Морган А., Джонс Н.Д., Несбитт А.М., Чаплин Л., Бодмер М.В., Emtage JS. N-конец домена Ch3 химерного IgG1 человека против HLA-DR необходим для связывания C1q, Fc гамма RI и Fc гамма RIII. Иммунология (1995) 86(2):319–24. [Статья бесплатно PMC] [PubMed] [Google Scholar]
133. Brekke OH, Michaelsen TE, Aase A, Sandin RH, Sandlie I.
134. Dall’Acqua WF, Cook KE, Damschroder MM, Woods RM, Wu H. Модуляция эффекторных функций IgG1 человека путем конструирования его шарнирной области. J Immunol (2006) 177(2):1129–38. 10.4049/jimmunol.177.2.1129 [PubMed] [CrossRef] [Google Scholar]
135. Dangl JL, Wensel TG, Morrison SL, Stryer L, Herzenberg LA, Oi VT. Сегментная гибкость и фиксация комплемента генетически сконструированных химерных антител человека, кролика и мыши. EMBO J (1988) 7 (7): 1989–94. [Бесплатная статья PMC] [PubMed] [Google Scholar]
136. Lu Y, Harding SE, Michaelsen TE, Longman E, Davis KG, Ortega A, et al. Конформация раствора дикого типа и мутантных иммуноглобулинов IgG3 и IgG4 с использованием кристаллогидродинамики: возможные последствия для активации комплемента.
137. Tan LK, Shopes RJ, Oi VT, Morrison SL. Влияние шарнирной области на активацию комплемента, связывание C1q и гибкость сегментов химерных иммуноглобулинов человека. Proc Natl Acad Sci U S A (1990) 87(1):162–6. 10.1073/pnas.87.1.162 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
138. Natsume A, In M, Takamura H, Nakagawa T, Shimizu Y, Kitajima K, et al. Сконструированы антитела смешанного изотипа IgG1/IgG3 с повышенной цитотоксической активностью. Рак Res (2008) 68 (10): 3863–72. 10.1158/0008-5472.CAN-07-6297 [PubMed] [CrossRef] [Google Scholar]
139. Aalberse RC, Schuurman J. IgG4 нарушает правила. Иммунология (2002) 105 (1): 9–19. 10.1046/j.0019-2805.2001.01341.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
140. Эйб Ю., Гор Дж., Брейсвелл Д.Г., Перкинс С.Дж., Долби П.А. Маскирование области Fc в IgG4 человека с помощью моделирования ограниченного рассеяния рентгеновских лучей: последствия для функции антител и терапии 1.
141. Van der Zee JS, Van Swieten P, Aalberse RC. Ингибирование активации комплемента антителами IgG4. Clin Exp Immunol (1986) 64(2):415–22. [Бесплатная статья PMC] [PubMed] [Google Scholar]
142. Saeland E, Vidarsson G, Leusen JH, Van Garderen E, Nahm MH, Vile-Weekhout H, et al. Центральная роль комплемента в пассивной защите человеческими антипневмококковыми антителами IgG1 и IgG2 у мышей. J Immunol (2003) 170(12):6158–64. 10.4049/jimmunol.170.12.6158 [PubMed] [CrossRef] [Google Scholar]
143. Vidarsson G, Jonsdottir I, Jonsson S, Valdimarsson H. Опсонизация и антитела к полисахаридам капсулы и клеточной стенки Streptococcus pneumoniae . J Infect Dis (1994) 170: 592–9. 10.1093/infdis/170.3.592 [PubMed] [CrossRef] [Google Scholar]
144. Ramsland PA, Farrugia W, Bradford TM, Sardjono CT, Esparon S, Trist HM, et al. Структурная основа распознавания Fc гаммаRIIa человеческим IgG и образования воспалительных сигнальных комплексов.
145. Guilliams M, Bruhns P, Saeys Y, Hammad H, Lambrecht BN. Функция рецепторов Fc в дендритных клетках и макрофагах. Nat Rev Immunol (2014) 14 (2): 94–108. 10.1038/nri3582 [PubMed] [CrossRef] [Google Scholar]
146. Ernst LK, van de Winkel JG, Chiu IM, Anderson CL. Три гена человеческого высокоаффинного Fc-рецептора для IgG (Fc гамма RI) кодируют четыре различных продукта транскрипции. Джей Биол Хим (1992) 267(22):15692–700. [PubMed] [Google Scholar]
147. Эрнст Л.К., Дюшемин А.М., Миллер К.Л., Андерсон К.Л. Молекулярная характеристика шести вариантов транскриптов Fcgamma рецептора класса I (CD64). Мол Иммунол (1998) 35 (14–15): 943–54. 10.1016/S0161-5890(98)00079-0 [PubMed] [CrossRef] [Google Scholar]
148. Hulett MD, Hogarth PM. Второй и третий внеклеточные домены FcgammaRI (CD64) обеспечивают уникальное высокоаффинное связывание IgG2a.
149. Кэнфилд С.М., Моррисон С.Л. Аффинность связывания человеческого IgG с его высокоаффинным Fc-рецептором определяется несколькими аминокислотами в домене Ch3 и модулируется шарнирной областью. J Exp Med (1991) 173(6):1483–91. 10.1084/jem.173.6.1483 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
150. Чаппел М.С., Изенман Д.Е., Эверетт М., Сюй Ю.Ю., Доррингтон К.Дж., Кляйн М.Х. Идентификация сайта связывания Fc-гамма-рецепторов класса I в IgG человека с помощью рекомбинантных гибридов IgG1/IgG2 и антител с точечной мутацией. Proc Natl Acad Sci U S A (1991) 88(20):9036–40. 10.1073/pnas.88.20.9036 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
151. Shields RL, Namenuk AK, Hong K, Meng YG, Rae J, Briggs J, et al. Картирование с высоким разрешением сайта связывания на IgG1 человека для Fc гамма RI, Fc гамма RII, Fc гамма RIII и FcRn и дизайн вариантов IgG1 с улучшенным связыванием с Fc гамма R.
152. Lund J, Winter G, Jones PT, Pound JD, Tanaka T, Walker MR, et al. Человеческий Fc гамма RI и Fc гамма RII взаимодействуют с разными, но перекрывающимися сайтами IgG человека. Джей Иммунол (1991) 147(8):2657–62. [PubMed] [Google Scholar]
153. Армор К.Л., Кларк М.Р., Хэдли А.Г., Уильямсон Л.М. Рекомбинантные молекулы IgG человека, лишенные активности связывания Fcgamma рецептора I и активации моноцитов. Eur J Immunol (1999) 29(8):2613–24. 10.1002/(SICI)1521-4141(199908)29:08<2613::AID-IMMU2613>3.0.CO;2-J [PubMed] [CrossRef] [Google Scholar]
154. Redpath S, Michaelsen TE, Sandlie Я, Кларк М.Р. Влияние длины шарнирной области на связывание IgG человека с Fcgamma-рецепторами человека. Хум Иммунол (1998) 59(11):720–7. 10.1016/S0198-8859(98)00075-5 [PubMed] [CrossRef] [Google Scholar]
155. Parren PW, Warmerdam PA, Boeije LC, Arts J, Westerdaal NA, Vlug A, et al.
156. Armor KL, van de Winkel JG, Williamson LM, Clark MR. Дифференциальное связывание человеческих рецепторов FcgammaRIIa и FcgammaRIIb человеческими IgG дикого типа и мутантными антителами. Мол Иммунол (2003) 40(9)): 585–93. 10.1016/j.molimm.2003.08.004 [PubMed] [CrossRef] [Google Scholar]
157. Вайнз Б.Д., Пауэлл М.С., Паррен П.В., Барнс Н., Хогарт П.М. Fc IgG содержит отдельные сайты связывания с рецептором Fc (FcR): рецепторы лейкоцитов Fc гамма RI и Fc гамма RIIa связываются с областью в Fc, отличной от той, которая распознается неонатальным FcR и белком A. J Immunol (2000) 164(10) : 5313–8. 10.4049/jimmunol.164.10.5313 [PubMed] [CrossRef] [Google Scholar]
158. van der Heijden J, Breunis WB, Geissler J, de Boer M, van den Berg TK, Kuijpers TW.
159. Breunis WB, van Mirre E, Bruin M, Geissler J, de BM, Peters M, et al. Изменение числа копий активирующего гена FCGR2C предрасполагает к идиопатической тромбоцитопенической пурпуре. Кровь (2008) 111 (3): 1029–38. 10.1182/blood-2007-03-079913 [PubMed] [CrossRef] [Google Scholar]
160. Li H, Sethuraman N, Stadheim TA, Zha D, Prinz B, Ballew N, et al. Оптимизация гуманизированных IgG в гликоинженерном Pichia pastoris . Nat Biotechnol (2006) 24(2):210–5. 10.1038/nbt1178 [PubMed] [CrossRef] [Google Scholar]
161. Siberil S, de Romeuf C, Bihoreau N, Fernandez N, Meterreau JL, Regenman A, et al. Выбор человеческого моноклонального антитела против RhD для терапевтического применения: влияние гликозилирования IgG на активирующие и ингибирующие функции Fc гамма R. Clin Immunol (2006) 118 (2–3): 170–9.
162. Huizinga TW, Kleijer M, Tetteroo PA, Roos D, von dem Borne AE. Двуаллельная нейтрофильная система Na-антигена связана с полиморфизмом фосфо-инозитол-связанного Fc гамма-рецептора III (CD16). Кровь (1990) 75(1):213–7. [PubMed] [Google Scholar]
163. Huizinga TW, Kerst M, Nuyens JH, Vlug A, von dem Borne AE, Roos D, et al. Характеристики связывания димерных комплексов подкласса IgG с нейтрофилами человека. J Immunol (1989) 142(7):2359–64. [PubMed] [Google Scholar]
164. Брамбелл Ф.В., Хеммингс В.А., Моррис И.Г. Теоретическая модель катаболизма гамма-глобулинов. Природа (1964) 203: 1352–4. 10.1038/2031352a0 [PubMed] [CrossRef] [Google Scholar]
165. Brambell FW. Передача иммунитета от матери к детенышу и катаболизм иммуноглобулинов. Ланцет (1966) 2(7473):1087–93. 10.1016/S0140-6736(66)92190-8 [PubMed] [CrossRef] [Google Scholar]
166. Ghetie V, Hubbard JG, Kim JK, Tsen MF, Lee Y, Ward ES.
167. Israel EJ, Wilsker DF, Hayes KC, Schoenfeld D, Simister NE. Повышенный клиренс IgG у мышей с дефицитом бета-2-микроглобулина: возможная защитная роль FcRn. Иммунология (1996) 89(4):573–8. 10.1046/j.1365-2567.1996.d01-775.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
168. Junghans RP, Anderson CL. Защитным рецептором катаболизма IgG является содержащий бета2-микроглобулин кишечный транспортный рецептор новорожденного. Proc Natl Acad Sci U S A (1996) 93(11):5512–6. 10.1073/pnas.93.11.5512 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
169. Roopenian DC, Christianson GJ, Sproule TJ, Brown AC, Akilesh S, Jung N, et al. Рецептор IgG, подобный MHC класса I, контролирует перинатальный транспорт IgG, гомеостаз IgG и судьбу препаратов, связанных с IgG-Fc.
170. Chaudhury C, Mehnaz S, Robinson JM, Hayton WL, Pearl DK, Roopenian DC, et al. Fc-рецептор IgG (FcRn), связанный с главным комплексом гистосовместимости, связывает альбумин и продлевает срок его жизни. J Biol Chem (2003) 197(3):315–22. 10.1084/jem.20021829 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
171. Gastinel LN, Simister NE, Bjorkman PJ. Экспрессия и кристаллизация растворимой и функциональной формы рецептора Fc, относящегося к молекулам гистосовместимости класса I. Proc Natl Acad Sci U S A (1992) 89(2):638–42. 10.1073/pnas.89.2.638 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
172. Einarsdottir H, Ji Y, Visser R, Mo C, Luo G, Scherjon S, et al. h535-содержащие аллотипы иммуноглобулина G3 эффективно транспортируются через плаценту человека: значение для опосредованных аллоантителами заболеваний новорожденных. Переливание (2014) 54 (3): 665–71.
173. Anderson CL, Chaudhury C, Kim J, Bronson CL, Wani MA, Mohanty S. Perspective – FcRn транспортирует альбумин: значение для иммунологии и медицины. Trends Immunol (2006) 27(7):343–8. 10.1016/j.it.2006.05.004 [PubMed] [CrossRef] [Google Scholar]
174. Borvak J, Richardson J, Medesan C, Antohe F, Radu C, Simionescu M, et al. Функциональная экспрессия рецептора, связанного с MHC класса I, FcRn, в эндотелиальных клетках мышей. Int Immunol (1998) 10(9):1289–98. 10.1093/intimm/10.9.1289 [PubMed] [CrossRef] [Google Scholar]
175. Акилеш С., Кристиансон Г.Дж., Рупениан Д.К., Шоу А.С. Неонатальная экспрессия FcR в клетках, происходящих из костного мозга, защищает сывороточный IgG от катаболизма. J Immunol (2007) 179(7):4580–8. 10.4049/иммунол.179.7.4580 [PubMed] [CrossRef] [Google Scholar]
176. Качковиц И., Червенак Дж., Эрдей А., Голдсби Р.А., Батлер Дж.Е. Недавние достижения в использовании сверхэкспрессии FcRn у трансгенных животных для преодоления препятствий стандартных технологий антител для улучшения выработки специфических антител.
177. Yoshida M, Claypool SM, Wagner JS, Mizoguchi E, Mizoguchi A, Roopenian DC, et al. Человеческий неонатальный Fc-рецептор опосредует транспорт IgG в просветные выделения для доставки антигенов к дендритным клеткам слизистой оболочки. Иммунитет (2004) 20(6):769–83. 10.1016/j.immuni.2004.05.007 [PubMed] [CrossRef] [Google Scholar]
178. Israel EJ, Patel VK, Taylor SF, Marshak-Rothstein A, Simister NE. Потребность в Fc-рецепторе, ассоциированном с бета-2-микроглобулином, для приобретения материнского IgG эмбрионами и новорожденными мышами. J Immunol (1995) 154(12):6246–51. [PubMed] [Google Scholar]
179. Israel EJ, Taylor S, Wu Z, Mizoguchi E, Blumberg RS, Bhan A, et al. Экспрессия неонатального рецептора Fc, FcRn, на эпителиальных клетках кишечника человека. Иммунология (1997) 92(1):69–74. 10.1046/j.1365-2567.1997.00326.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
180.
181. Spiekermann GM, Finn PW, Ward ES, Dumont J, Dickinson BL, Blumberg RS, et al. Рецептор-опосредованный транспорт иммуноглобулина G через слизистые барьеры во взрослой жизни: функциональная экспрессия FcRn в легких млекопитающих. J Exp Med (2002) 196(3):303–10. 10.1084/jem.20020400 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
182. Claypool SM, Dickinson BL, Wagner JS, Johansen FE, Venu N, Borawski JA, et al. Двунаправленный трансэпителиальный транспорт IgG сильно поляризованным базолатеральным мембранным Fcgamma-рецептором. Mol Biol Cell (2004) 15(4):1746–59. 10.1091/mbc.E03-11-0832 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
183. Dickinson BL, Badizadegan K, Wu Z, Ahouse JC, Zhu X, Simister NE, et al.
184. Shah U, Dickinson BL, Blumberg RS, Simister NE, Lencer WI, Walker WA. Распределение Fc-рецептора IgG, FcRn, в кишечнике плода человека. Педиатр Рез (2003) 53 (2): 295–301. 10.1203/01.PDR.0000047663.81816.E3 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
185. Карам-Салас Н., Буало Э., Фаррингтон Г.К., Гарбер Э., Брюнет Э., Абулроб А. и др. . Методы in vitro и in vivo для оценки FcRn-опосредованного обратного трансцитоза через гематоэнцефалический барьер. Methods Mol Biol (2011) 763:383–401. 10.1007/978-1-61779-191-8_26 [PubMed] [CrossRef] [Google Scholar]
186. Роберт-Гурофф М. Поверхности IgG как важный компонент защиты слизистой оболочки. Nat Med (2000) 6 (2): 129–30. 10.1038/72206 [PubMed] [CrossRef] [Google Scholar]
187.
188. Horton RE, Vidarsson G. Антитела и их рецепторы: различные потенциальные роли в защите слизистой оболочки. Фронт Иммунол (2013) 4:200. 10.3389/fimmu.2013.00200 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
189. Zhu X, Meng G, Dickinson BL, Li X, Mizoguchi E, Miao L, et al. Связанный с МНС класса I неонатальный Fc-рецептор для IgG функционально экспрессируется в моноцитах, кишечных макрофагах и дендритных клетках. J Immunol (2001) 166(5):3266–76. 10.4049/jimmunol.166.5.3266 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
190. Cervenak J, Bender B, Schneider Z, Magna M, Carstea BV, Liliom K, et al. Сверхэкспрессия неонатального FcR усиливает гуморальный иммунный ответ у трансгенных мышей.
191. Vidarsson G, Stemerding AM, Stapleton NM, Spliethoff SE, Janssen H, Rebers FE, et al. FcRn: рецептор IgG на фагоцитах с новой ролью в фагоцитозе. Кровь (2006) 108 (10): 3573–9. 10.1182/blood-2006-05-024539 [PubMed] [CrossRef] [Google Scholar]
192. Baker K, Qiao SW, Kuo TT, Aveson VG, Platzer B, Andersen JT, et al. Неонатальный Fc-рецептор для IgG (FcRn) регулирует перекрестную презентацию иммунных комплексов IgG дендритными клетками CD8-CD11b+. Proc Natl Acad Sci U S A (2011) 108(24):9927–32. 10.1073/pnas.10108 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
193. Mi W, Wanjie S, Lo ST, Gan Z, Pickl-Herk B, Ober RJ, et al. Нацеливание на неонатальный Fc-рецептор для доставки антигена с использованием сконструированных Fc-фрагментов. J Immunol (2008) 181(11):7550. 10.4049/jиммунol.181.11.7550 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
194.
195. Heijnen I, van Vugt MJ, Fanger N, Graziano RF, de Wit TP, Hofhuis FM, et al. Нацеливание антигена на миелоид-специфический человеческий Fc гамма RI/CD64 вызывает усиленный ответ антител у трансгенных мышей. Джей Клин Инвест (1996) 97(2):331–8. 10.1172/JCI118420 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
196. Ehrhardt GR, Davis RS, Hsu JT, Leu CM, Ehrhardt A, Cooper MD. Ингибирующий потенциал гомолога 4 рецептора Fc на В-клетках памяти. Proc Natl Acad Sci U S A (2003) 100(23):13489–94. 10.1073/pnas.1935944100 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
197. Haga CL, Ehrhardt GR, Boohaker RJ, Davis RS, Cooper MD. Fc-рецептор-подобный 5 ингибирует активацию В-клеток посредством рекрутирования тирозинфосфатазы SHP-1.
198. Polson AG, Zheng B, Elkins K, Chang W, Du C, Dowd P, et al. Характер экспрессии рецепторов FcRH/IRTA человека в нормальных тканях и при В-хроническом лимфоцитарном лейкозе. Int Immunol (2006) 18(9):1363–73. 10.1093/intimm/dxl069 [PubMed] [CrossRef] [Google Scholar]
199. Ehrhardt GR, Hsu JT, Gartland L, Leu CM, Zhang S, Davis RS, et al. Экспрессия иммунорегуляторной молекулы FcRh5 определяет характерную тканевую популяцию В-клеток памяти. J Exp Med (2005) 202 (6): 783–91. 10.1084/jem.20050879 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
200. Frank MB, Itoh K, Fujisaku A, Pontarotti P, Mattei MG, Neas BR. Картирование гена аутоантигена Ro/SSA человека массой 52 кДа на хромосому 11 человека и его полиморфизмы. Am J Hum Genet (1993) 52 (1): 183–91. [Бесплатная статья PMC] [PubMed] [Google Scholar]
201. Yang YS, Yang MC, Wang B, Weissler JC.
202. Rhodes D, Trowsdale J. TRIM21 представляет собой тримерный белок, который связывает Fc IgG через домен B30.2. Мол Иммунол (2007) 44(9):2406–14. 10.1016/j.molimm.2006.10.013 [PubMed] [CrossRef] [Google Scholar]
203. Vaysburd M, Watkinson RE, Cooper H, Reed M, O’Connell K, Smith J, et al. Рецептор внутриклеточного антитела TRIM21 предотвращает смертельную вирусную инфекцию. Proc Natl Acad Sci USA (2013) 110(30):12397–401. 10.1073/пнас.1301
0 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
204. Watkinson RE, Tam JC, Vaysburd MJ, James LC. Одновременная нейтрализация и обнаружение врожденным иммунитетом реплицирующегося вируса с помощью TRIM21. Дж. Вирол (2013) 87 (13): 7309–13. 10.1128/JVI.00647-13 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
205.
206. Mallery DL, McEwan W, Bidgood SR, Towers GJ, Johnson CM, James LC. Антитела опосредуют внутриклеточный иммунитет через тройной мотив, содержащий 21 (TRIM21). Proc Natl Acad Sci U S A (2010) 107(46):19985–90. 10.1073/pnas.1014074107 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
207. McEwan W, Mallery DL, Rhodes D, Trowsdale J, James LC. Внутриклеточный иммунитет, опосредованный антителами, и роль TRIM21. Биоэссеи (2011) 33 (11): 803–9. 10.1002/bies.201100093 [PubMed] [CrossRef] [Google Scholar]
208. McEwan W, Tam JC, Watkinson RE, Bidgood SR, Mallery DL, James LC. Внутриклеточные патогены, связанные с антителами, стимулируют передачу иммунных сигналов через Fc-рецептор TRIM21. Nat Immunol (2013) 14(4):327–36. 10.
209. Anthony RM, Kobayashi T, Wermeling F, Ravetch JV. Внутривенный гаммаглобулин подавляет воспаление посредством нового пути T (H) 2 . Природа (2011) 475 (7354): 110–3. 10.1038/nature10134 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
210. Энтони Р.М., Вермелинг Ф., Карлссон М.С., Раветч СП. Идентификация рецептора, необходимого для противовоспалительной активности ВВИГ. Proc Natl Acad Sci U S A (2008) 105(50):19571–8. 10.1073/pnas.0810163105 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
211. Massoud AH, Yona M, Xue D, Chouiali F, Alturaihi H, Ablona A, et al. Иммунорецептор дендритных клеток: новый рецептор для внутривенного иммуноглобулина опосредует индукцию регуляторных Т-клеток. J Allergy Clin Immunol (2014) 133(3):853–63. 10.1016/j.jaci.2013.09.029 [PubMed] [CrossRef] [Google Scholar]
212. Yu X, Vasiljevic S, Mitchell D, Crispin M, Scanlan CN.
213. von Gunten S, Schaub A, Vogel M, Stadler BM, Miescher S, Simon HU. Иммунологические и функциональные доказательства наличия аутоантител против Siglec-9 в препаратах внутривенного иммуноглобулина. Кровь (2006) 108 (13): 4255–9. 10.1182/blood-2006-05-021568 [PubMed] [CrossRef] [Google Scholar]
214. von Gunten S, Vogel M, Schaub A, Stadler BM, Miescher S, Crocker PR, et al. Внутривенные препараты иммуноглобулина содержат аутоантитела против Siglec-8. J Allergy Clin Immunol (2007) 119(4):1005–11. 10.1016/j.jaci.2007.01.023 [PubMed] [CrossRef] [Google Scholar]
215. Seite JF, Cornec D, Renaudineau Y, Youinou P, Mageed RA, Hillion S. IVIg модулирует передачу сигналов BCR через CD22 и способствует апоптозу. в зрелых В-лимфоцитах человека.
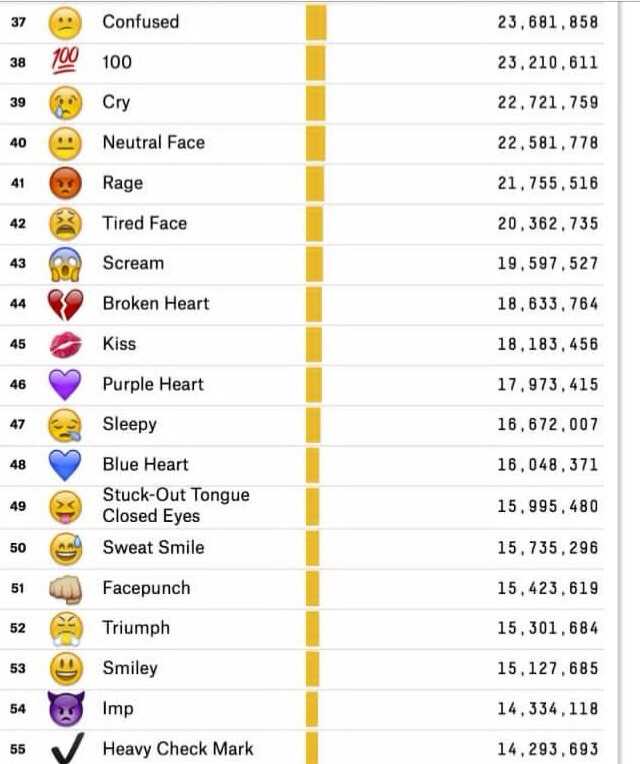 гора!
гора! Типичный формат гоночного уик-энда выглядит примерно так:
Типичный формат гоночного уик-энда выглядит примерно так:
 Вот несколько известных рас:
Вот несколько известных рас:
 Формат простой: спортсмены взбираются по крутым склонам на высоту 1000 м по трассе, обычно не превышающей 5 км.
Формат простой: спортсмены взбираются по крутым склонам на высоту 1000 м по трассе, обычно не превышающей 5 км.
 Изучение топографической карты или профиля высот, показывающих относительную крутизну местности, даст вам представление о том, где вы сможете бежать, а где придется идти пешком.
Изучение топографической карты или профиля высот, показывающих относительную крутизну местности, даст вам представление о том, где вы сможете бежать, а где придется идти пешком. com
com Двигайтесь небольшими шагами
Двигайтесь небольшими шагами
 Привнесите немного альпийской ва-ва-вумы в свой первый ВКонтакте, вооружив своих чирлидеров подходящим инструментом для работы!
Привнесите немного альпийской ва-ва-вумы в свой первый ВКонтакте, вооружив своих чирлидеров подходящим инструментом для работы! Ассоциированные транспозазы (CAST) обеспечивают рекомбинационно-независимые вставки ДНК с несколькими тысячами оснований в запрограммированных РНК местах генома. CAST типа V-K предлагают явные технологические преимущества по сравнению с CAST типа I, учитывая их меньший размер кода, меньшее количество компонентов и однонаправленные вставки. Однако полезность CAST типа V-K затруднена механизмом репликативной транспозиции, который приводит к смеси желаемых простых вставок грузов и нежелательных продуктов коинтеграции плазмид. Здесь мы преодолеваем это ограничение, разрабатывая новые CAST с значительно улучшенной чистотой продукта. Для этого мы компенсируем отсутствие субъединицы TnsA в множественных CAST типа V-K, разрабатывая систему CAST с большой последовательностью, интегрирующую самонаводящиеся эндонуклеазы, или систему HELIX. HELIX использует самонаводящуюся эндонуклеазу (nHE), слитую с TnsB, для восстановления 5-дюймовой способности к надрезу, необходимой для двойного разрыва донора ДНК.
Ассоциированные транспозазы (CAST) обеспечивают рекомбинационно-независимые вставки ДНК с несколькими тысячами оснований в запрограммированных РНК местах генома. CAST типа V-K предлагают явные технологические преимущества по сравнению с CAST типа I, учитывая их меньший размер кода, меньшее количество компонентов и однонаправленные вставки. Однако полезность CAST типа V-K затруднена механизмом репликативной транспозиции, который приводит к смеси желаемых простых вставок грузов и нежелательных продуктов коинтеграции плазмид. Здесь мы преодолеваем это ограничение, разрабатывая новые CAST с значительно улучшенной чистотой продукта. Для этого мы компенсируем отсутствие субъединицы TnsA в множественных CAST типа V-K, разрабатывая систему CAST с большой последовательностью, интегрирующую самонаводящиеся эндонуклеазы, или систему HELIX. HELIX использует самонаводящуюся эндонуклеазу (nHE), слитую с TnsB, для восстановления 5-дюймовой способности к надрезу, необходимой для двойного разрыва донора ДНК. Используя отличительные особенности систем типа V-K и типа I, HELIX позволяет вставлять ДНК методом вырезания и вставки с помощью до 9Чистота продукта простой вставки составляет 9,3 % при сохранении надежной эффективности интеграции с геномными мишенями. Кроме того, мы демонстрируем универсальность этого подхода, создавая системы HELIX для других ортологов CAST. Мы также устанавливаем возможность создания минимальной 3-компонентной HELIX, упрощая количество белков, которые должны быть экспрессированы. Вместе HELIX оптимизирует и улучшает применение технологий транспозиции на основе CRISPR, устраняя барьеры для эффективных и специфических вставок ДНК, управляемых РНК.
Используя отличительные особенности систем типа V-K и типа I, HELIX позволяет вставлять ДНК методом вырезания и вставки с помощью до 9Чистота продукта простой вставки составляет 9,3 % при сохранении надежной эффективности интеграции с геномными мишенями. Кроме того, мы демонстрируем универсальность этого подхода, создавая системы HELIX для других ортологов CAST. Мы также устанавливаем возможность создания минимальной 3-компонентной HELIX, упрощая количество белков, которые должны быть экспрессированы. Вместе HELIX оптимизирует и улучшает применение технологий транспозиции на основе CRISPR, устраняя барьеры для эффективных и специфических вставок ДНК, управляемых РНК. DSB и механизм гомологичной рекомбинации хозяина. Кроме того, системы рекомбинации у бактерий имеют низкую эффективность 11 без коинтеграции селективного маркера 12 или контрселекции CRISPR-Cas 13 . CRISPR-ассоциированные транспозазы (CAST) представляют собой многообещающий новый подход к программируемым, независимым от рекомбинации вставкам ДНК посредством взаимодействия между механизмом транспозазы и эффектором(ами) CRISPR для прямой транспозиции, управляемой РНК 14-16 .
DSB и механизм гомологичной рекомбинации хозяина. Кроме того, системы рекомбинации у бактерий имеют низкую эффективность 11 без коинтеграции селективного маркера 12 или контрселекции CRISPR-Cas 13 . CRISPR-ассоциированные транспозазы (CAST) представляют собой многообещающий новый подход к программируемым, независимым от рекомбинации вставкам ДНК посредством взаимодействия между механизмом транспозазы и эффектором(ами) CRISPR для прямой транспозиции, управляемой РНК 14-16 . Кроме того, склонность некоторых систем типа I к двунаправленным вставкам приводит к нежелательной примеси редактирования 15 ( рис. 1а ). Для сравнения, CAST типа V-K более компактны с точки зрения размера кода, содержат только четыре основных компонента и обеспечивают полную или почти полную однонаправленную вставку. Однако CAST типа V-K приводят к проблемной смеси продуктов простой вставки и коинтеграции генов, последняя из которых состоит из дупликации груза и полной вставки остова плазмиды 4,6 ( рис. 1b ), и в целом имеют более низкую специфичность 14,16,17 . Для приложений редактирования генома идеальная технология встраивания ДНК позволила бы создавать программируемые, однонаправленные, независимые от рекомбинации и простые генные продукты вставки с минимальными компонентами и оборудованием минимального размера. Поэтому мы стремились разработать инженерную технологию CAST, сочетающую простоту и предсказуемость ориентации систем типа V-K с чистотой продукта систем типа I.
Кроме того, склонность некоторых систем типа I к двунаправленным вставкам приводит к нежелательной примеси редактирования 15 ( рис. 1а ). Для сравнения, CAST типа V-K более компактны с точки зрения размера кода, содержат только четыре основных компонента и обеспечивают полную или почти полную однонаправленную вставку. Однако CAST типа V-K приводят к проблемной смеси продуктов простой вставки и коинтеграции генов, последняя из которых состоит из дупликации груза и полной вставки остова плазмиды 4,6 ( рис. 1b ), и в целом имеют более низкую специфичность 14,16,17 . Для приложений редактирования генома идеальная технология встраивания ДНК позволила бы создавать программируемые, однонаправленные, независимые от рекомбинации и простые генные продукты вставки с минимальными компонентами и оборудованием минимального размера. Поэтому мы стремились разработать инженерную технологию CAST, сочетающую простоту и предсказуемость ориентации систем типа V-K с чистотой продукта систем типа I.
 h , Распределение длины считывания при использовании ShCAST и ShHELIX с sgRNA, нацеленной на TS1 на pTarget, из данных секвенирования длительного считывания. Верхняя правая панель представляет собой увеличенное изображение пика длины считывания ~8000 п.н. и , Сравнение пропорций продуктов простой вставки и коинтеграции транспонированных продуктов для конструкций ShCAST и ShHELIX при использовании pDonor с сайтами I-AniI в 14 п.н. последовательность чтения. й , k , Чистота продукта транспозиции ( панель j ) и КОЕ ( панель k ) при использовании сайта Lib4 I-AniI на pDonor (с расстоянием 14 п.н. между сайтами Lib4 и LE/RE), которые ранее было показано, что он увеличивает аффинность I-AniI дикого типа в 5 раз. Для панелей f и k , среднее, стандартное отклонение и отдельные точки данных показаны для n = 3. TSD, дублирование целевого сайта; LE и RE — левый и правый концы транспозона соответственно; sgRNA, одиночная направляющая РНК; ddPCR, капельная цифровая ПЦР.
h , Распределение длины считывания при использовании ShCAST и ShHELIX с sgRNA, нацеленной на TS1 на pTarget, из данных секвенирования длительного считывания. Верхняя правая панель представляет собой увеличенное изображение пика длины считывания ~8000 п.н. и , Сравнение пропорций продуктов простой вставки и коинтеграции транспонированных продуктов для конструкций ShCAST и ShHELIX при использовании pDonor с сайтами I-AniI в 14 п.н. последовательность чтения. й , k , Чистота продукта транспозиции ( панель j ) и КОЕ ( панель k ) при использовании сайта Lib4 I-AniI на pDonor (с расстоянием 14 п.н. между сайтами Lib4 и LE/RE), которые ранее было показано, что он увеличивает аффинность I-AniI дикого типа в 5 раз. Для панелей f и k , среднее, стандартное отклонение и отдельные точки данных показаны для n = 3. TSD, дублирование целевого сайта; LE и RE — левый и правый концы транспозона соответственно; sgRNA, одиночная направляющая РНК; ddPCR, капельная цифровая ПЦР.
 1c). ). Идеальная никаза должна быть небольшой (чтобы добавить в систему минимальный размер кодирования), иметь предсказуемый сайт разрыва и предпочтительность нити, обладать высокой специфичностью для предотвращения непреднамеренного разрыва по мишени или вне ее и функционировать в различных организмах для разработки последующих инструментов. и приложение. Потенциальные варианты включают эндонуклеазы рестрикции 22 , фрагментированные варианты Cas 9,23,24 , каталитическая часть или полный фермент TnsA из CAST типа I или транспозонов Tn7 25 , или фаговые эндонуклеазы HNH 26 . Однако у каждого из них есть недостатки, касающиеся размера, специфичности последовательности и цепи и / или необходимых белок-белковых взаимодействий.
1c). ). Идеальная никаза должна быть небольшой (чтобы добавить в систему минимальный размер кодирования), иметь предсказуемый сайт разрыва и предпочтительность нити, обладать высокой специфичностью для предотвращения непреднамеренного разрыва по мишени или вне ее и функционировать в различных организмах для разработки последующих инструментов. и приложение. Потенциальные варианты включают эндонуклеазы рестрикции 22 , фрагментированные варианты Cas 9,23,24 , каталитическая часть или полный фермент TnsA из CAST типа I или транспозонов Tn7 25 , или фаговые эндонуклеазы HNH 26 . Однако у каждого из них есть недостатки, касающиеся размера, специфичности последовательности и цепи и / или необходимых белок-белковых взаимодействий. LAGLIDADG HEs (LHE) были использованы для редактирования генома в бактериальных и человеческих клетках из-за их специфичности и их умеренной перепрограммируемости с помощью белковой инженерии или химерной сборки 27 . LHE из Aspergillus nidulans (I-AniI) имеет небольшую кодирующую последовательность (254 аминокислоты), расщепляет асимметричную последовательность ДНК-мишень длиной 19 п. 28 (нАниИ). Кроме того, было показано, что гиперактивный вариант I-AniI, названный Y2 I-AniI, обладает в 9 раз большей аффинностью к родственному ему сайту-мишени 29 . Поэтому мы предположили, что либо nAniI, либо Y2 nAniI могут быть слиты с TnsB, чтобы сделать возможным двойное никирование на донорской плазмиде, необходимое для вставки ДНК путем вырезания и вставки с помощью CAST типа V-K (9).0009 Рис. 1с ). Вместе слияние nHE с TnsB вместе с остальными компонентами CAST образует комплекс CAST с большой последовательностью с помощью HE, или HELIX.
LAGLIDADG HEs (LHE) были использованы для редактирования генома в бактериальных и человеческих клетках из-за их специфичности и их умеренной перепрограммируемости с помощью белковой инженерии или химерной сборки 27 . LHE из Aspergillus nidulans (I-AniI) имеет небольшую кодирующую последовательность (254 аминокислоты), расщепляет асимметричную последовательность ДНК-мишень длиной 19 п. 28 (нАниИ). Кроме того, было показано, что гиперактивный вариант I-AniI, названный Y2 I-AniI, обладает в 9 раз большей аффинностью к родственному ему сайту-мишени 29 . Поэтому мы предположили, что либо nAniI, либо Y2 nAniI могут быть слиты с TnsB, чтобы сделать возможным двойное никирование на донорской плазмиде, необходимое для вставки ДНК путем вырезания и вставки с помощью CAST типа V-K (9).0009 Рис. 1с ). Вместе слияние nHE с TnsB вместе с остальными компонентами CAST образует комплекс CAST с большой последовательностью с помощью HE, или HELIX.
 Интересно, что слияние nAniI с N-концом TnsB поддерживало РНК-управляемую вставку ДНК (9).0009 Рис. 1e ). С-концевые слияния nAniI с TnsB не приводили к транспозиции, указывая на то, что С-концевой взаимодействующий с TnsC домен TnsB в меньшей степени подходит для слитых белков 30 .
Интересно, что слияние nAniI с N-концом TnsB поддерживало РНК-управляемую вставку ДНК (9).0009 Рис. 1e ). С-концевые слияния nAniI с TnsB не приводили к транспозиции, указывая на то, что С-концевой взаимодействующий с TnsC домен TnsB в меньшей степени подходит для слитых белков 30 . н. обеспечивал наивысшую интеграцию (, рис. 1f, ). Удивительно, но ShCAST также продемонстрировал переменную эффективность интеграции в зависимости от расстояния между сайтом I-AniI и LE/RE (где, в отличие от ShHELIX, сайт I-AniI не играет роли в транспозиции). Интервалы в 4–12 п.н. на pDonor приводили к значительно более высокой эффективности вставки, чем pDonor без сайтов I-AniI (9).0009 доп. рис. 1b ). Изменение положения сайта I-AniI модифицирует последовательность, непосредственно примыкающую к LE/RE на pDonor, что позволяет предположить, что состав фланкирующей последовательности, особенно первые 12 п.н., может быть важным фактором, определяющим эффективность интеграции ( Sup. Fig. 1б ). Отдельно мы также провели эксперименты по интеграции с использованием Y2 nAniI, слитого с TnsB (Y2 ShHELIX), и наблюдали значительно меньшее количество колоний с пиковым числом с интервалом 14 п.н. ( Sup. Fig. 3a 9).0010 и Доп. Примечание.
н. обеспечивал наивысшую интеграцию (, рис. 1f, ). Удивительно, но ShCAST также продемонстрировал переменную эффективность интеграции в зависимости от расстояния между сайтом I-AniI и LE/RE (где, в отличие от ShHELIX, сайт I-AniI не играет роли в транспозиции). Интервалы в 4–12 п.н. на pDonor приводили к значительно более высокой эффективности вставки, чем pDonor без сайтов I-AniI (9).0009 доп. рис. 1b ). Изменение положения сайта I-AniI модифицирует последовательность, непосредственно примыкающую к LE/RE на pDonor, что позволяет предположить, что состав фланкирующей последовательности, особенно первые 12 п.н., может быть важным фактором, определяющим эффективность интеграции ( Sup. Fig. 1б ). Отдельно мы также провели эксперименты по интеграции с использованием Y2 nAniI, слитого с TnsB (Y2 ShHELIX), и наблюдали значительно меньшее количество колоний с пиковым числом с интервалом 14 п.н. ( Sup. Fig. 3a 9).0010 и Доп. Примечание. 1 ). Для последующих экспериментов использовали конструкции HELIX со слияниями nAniI-TnsB и pDonors с 14 п.н. между сайтами I-AniI и LE/RE.
1 ). Для последующих экспериментов использовали конструкции HELIX со слияниями nAniI-TnsB и pDonors с 14 п.н. между сайтами I-AniI и LE/RE. Кроме того, мы не наблюдали сайты I-AniI в считываниях продукта вставки, предполагая, что 5′-фланец, содержащий эти последовательности, удаляется во время разрешения HELIX-опосредованной транспозиции. Мы также выполнили секвенирование с длительным считыванием с помощью Y2 ShHELIX и аналогичным образом наблюдали резкое увеличение чистоты продукта простой вставки (9).0009 доп. Рис. 3б-д ).
Кроме того, мы не наблюдали сайты I-AniI в считываниях продукта вставки, предполагая, что 5′-фланец, содержащий эти последовательности, удаляется во время разрешения HELIX-опосредованной транспозиции. Мы также выполнили секвенирование с длительным считыванием с помощью Y2 ShHELIX и аналогичным образом наблюдали резкое увеличение чистоты продукта простой вставки (9).0009 доп. Рис. 3б-д ). Fig. 4b ). Промежуточное образование шпильки или промежуточное расщепление Шапиро, индуцированное повреждением (в дополнение к возможности низкоуровневой DSB-опосредованной эксцизии груза), может происходить, что приводит к простым продуктам вставки ( Sup. Fig. 4c ). Наконец, использование сайта I-AniI «Lib4» на pDonor-14, как было обнаружено ранее, увеличивает аффинность I-AniI дикого типа в 5 раз 31 , еще больше снизил коинтеграты относительно ShHELIX до 0,23% всех продуктов транспозиции (для 84-кратного снижения коинтегратов по сравнению с ShCAST) ( рис. 1j ). Однако это улучшение чистоты продукта также сопровождалось снижением КОЕ (, примечание 1, и , рис. 1k, ). В целом, ShHELIX в сочетании с сайтом I-AniI, ориентированным на pDonor для создания 5-дюймового разрыва, демонстрирует наиболее заметное увеличение доли простой вставки до процента коинтеграции, что приводит к почти идеальной чистоте продукта на плазмидной мишени.
Fig. 4b ). Промежуточное образование шпильки или промежуточное расщепление Шапиро, индуцированное повреждением (в дополнение к возможности низкоуровневой DSB-опосредованной эксцизии груза), может происходить, что приводит к простым продуктам вставки ( Sup. Fig. 4c ). Наконец, использование сайта I-AniI «Lib4» на pDonor-14, как было обнаружено ранее, увеличивает аффинность I-AniI дикого типа в 5 раз 31 , еще больше снизил коинтеграты относительно ShHELIX до 0,23% всех продуктов транспозиции (для 84-кратного снижения коинтегратов по сравнению с ShCAST) ( рис. 1j ). Однако это улучшение чистоты продукта также сопровождалось снижением КОЕ (, примечание 1, и , рис. 1k, ). В целом, ShHELIX в сочетании с сайтом I-AniI, ориентированным на pDonor для создания 5-дюймового разрыва, демонстрирует наиболее заметное увеличение доли простой вставки до процента коинтеграции, что приводит к почти идеальной чистоте продукта на плазмидной мишени.
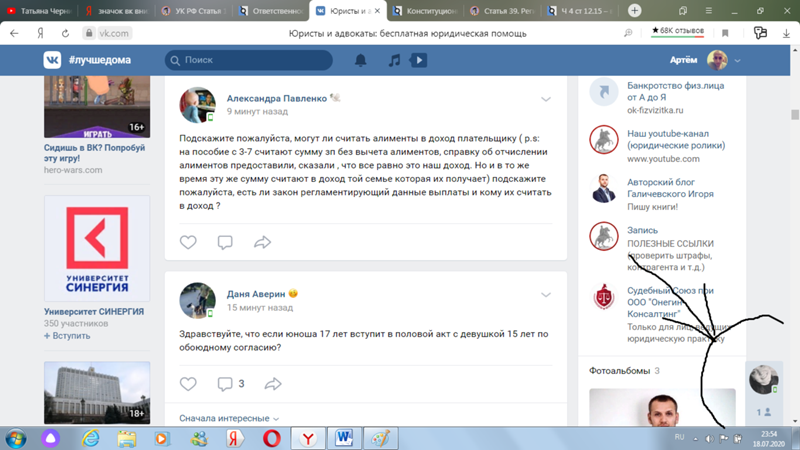 2c, ).
2c, ). Транспонированные продукты были обогащены перед секвенированием с помощью целевого обогащения Cas9. f , Распределение длины считывания продуктов транспозиции при использовании ShCAST и ShHELIX на геномном целевом сайте 2 (TS2) из данных секвенирования длительного считывания. Верхняя правая панель представляет собой увеличенное представление пика длины считывания ~8200 п.н. г , Сравнение пропорций продуктов простой вставки и коинтеграции при TS2 для ShCAST и ShHELIX, оцениваемых с помощью секвенирования с длительным чтением. h , Эффективность интеграции с ShHELIX и sgRNA, нацеленной на TS5, при использовании pDonors, кодирующих грузы различных размеров. Интеграцию оценивали с помощью ddPCR. Для панелей b, d и h , среднее значение, стандартное отклонение и отдельные точки данных показаны для n = 3. LE и RE, левый и правый концы транспозона, соответственно; sgRNA, одиночная направляющая РНК; ddPCR, капельная цифровая ПЦР.
Транспонированные продукты были обогащены перед секвенированием с помощью целевого обогащения Cas9. f , Распределение длины считывания продуктов транспозиции при использовании ShCAST и ShHELIX на геномном целевом сайте 2 (TS2) из данных секвенирования длительного считывания. Верхняя правая панель представляет собой увеличенное представление пика длины считывания ~8200 п.н. г , Сравнение пропорций продуктов простой вставки и коинтеграции при TS2 для ShCAST и ShHELIX, оцениваемых с помощью секвенирования с длительным чтением. h , Эффективность интеграции с ShHELIX и sgRNA, нацеленной на TS5, при использовании pDonors, кодирующих грузы различных размеров. Интеграцию оценивали с помощью ddPCR. Для панелей b, d и h , среднее значение, стандартное отклонение и отдельные точки данных показаны для n = 3. LE и RE, левый и правый концы транспозона, соответственно; sgRNA, одиночная направляющая РНК; ddPCR, капельная цифровая ПЦР.
 С ShHELIX мы наблюдали только 3,11% коинтегратов, что в 17 раз меньше по сравнению с ShCAST (, рис. 2e-g, ).
С ShHELIX мы наблюдали только 3,11% коинтегратов, что в 17 раз меньше по сравнению с ShCAST (, рис. 2e-g, ). . Для этого мы разработали трехкомпонентные системы, исследуя слияния различных субъединиц ShCAST. Мы исследовали слияние одного или двух мономеров TniQ с N- или С-концом Cas12k или слияние одного мономера TnsC с любым концом Cas12k (9).0009 Рис. 3а ). Слияние либо TniQ, либо TnsC с N-концом Cas12k существенно снижает эффективность интеграции (, рис. 3b, ). С-концевые слияния TniQ с Cas12k сохраняли эффективность интеграции до 62,3% и 69,6% по сравнению с неконденсированным ShCAST, когда один или два мономера TniQ были слиты соответственно (, рис. 3b, и , суп., рис. 6a, ). . Мы также наблюдали, что слияние мономера TnsC с С-концом Cas12k сохраняет относительно высокую эффективность интеграции (9).0009 Рис. 3b ). Учитывая предполагаемую критическую роль образования филаментов TnsC для транспозиции 30,33-35 , мы предполагаем, что филаментация все еще может происходить, несмотря на слияние Cas12k, или что только одного мономера достаточно для обеспечения транспозиции посредством неканонического механизма ( Приложение 3 ).
. Для этого мы разработали трехкомпонентные системы, исследуя слияния различных субъединиц ShCAST. Мы исследовали слияние одного или двух мономеров TniQ с N- или С-концом Cas12k или слияние одного мономера TnsC с любым концом Cas12k (9).0009 Рис. 3а ). Слияние либо TniQ, либо TnsC с N-концом Cas12k существенно снижает эффективность интеграции (, рис. 3b, ). С-концевые слияния TniQ с Cas12k сохраняли эффективность интеграции до 62,3% и 69,6% по сравнению с неконденсированным ShCAST, когда один или два мономера TniQ были слиты соответственно (, рис. 3b, и , суп., рис. 6a, ). . Мы также наблюдали, что слияние мономера TnsC с С-концом Cas12k сохраняет относительно высокую эффективность интеграции (9).0009 Рис. 3b ). Учитывая предполагаемую критическую роль образования филаментов TnsC для транспозиции 30,33-35 , мы предполагаем, что филаментация все еще может происходить, несмотря на слияние Cas12k, или что только одного мономера достаточно для обеспечения транспозиции посредством неканонического механизма ( Приложение 3 ).
 3c, ). Мы выполнили преобразования с использованием слияний Cas12k-TniQ, Cas12k-TniQ-TniQ или Cas12k-TnsC вместе с оставшимися модулями HELIX и оценили эффективность интеграции. Мы наблюдали РНК-управляемую интеграцию между 22-36% при TS2 и 15-49% при TS5 в зависимости от используемой архитектуры слияния (, рис. 3d, и , см. рис. 6b, 9).0010). Секвенирование ампликонов следующего поколения показало, что 4- и 3-компонентные системы ShCAST и ShHELIX демонстрируют один и тот же профиль расстояния вставки PAM-LE (, суп. рис. 7, ), показывая, что слияния nAniI-TnsB и слияния TniQ или TnsC Cas12k, отдельно или в сочетании, не изменяет это свойство. Эти результаты демонстрируют возможность создания систем HELIX с меньшим количеством субъединиц путем слияния компонентов и подчеркивают возможности белковой инженерии для создания трехкомпонентных версий, демонстрирующих сопоставимую эффективность интеграции.
3c, ). Мы выполнили преобразования с использованием слияний Cas12k-TniQ, Cas12k-TniQ-TniQ или Cas12k-TnsC вместе с оставшимися модулями HELIX и оценили эффективность интеграции. Мы наблюдали РНК-управляемую интеграцию между 22-36% при TS2 и 15-49% при TS5 в зависимости от используемой архитектуры слияния (, рис. 3d, и , см. рис. 6b, 9).0010). Секвенирование ампликонов следующего поколения показало, что 4- и 3-компонентные системы ShCAST и ShHELIX демонстрируют один и тот же профиль расстояния вставки PAM-LE (, суп. рис. 7, ), показывая, что слияния nAniI-TnsB и слияния TniQ или TnsC Cas12k, отдельно или в сочетании, не изменяет это свойство. Эти результаты демонстрируют возможность создания систем HELIX с меньшим количеством субъединиц путем слияния компонентов и подчеркивают возможности белковой инженерии для создания трехкомпонентных версий, демонстрирующих сопоставимую эффективность интеграции. Это наблюдение поддерживает эволюционную гипотезу о том, что Tn5053-подобный транспозон, содержащий TnsB, TnsC и TniQ, но не TnsA, ассимилировал и перепрофилировал эту систему CRISPR 19 . Следовательно, ожидается, что все CAST типа V-K будут действовать посредством репликативной транспозиции, приводя к существенной доле нежелательных продуктов коинтеграции. Таким образом, мы исследовали HELIX в качестве универсального подхода, позволяющего вставлять ДНК методом вырезания и вставки с другими V-K CAST различных типов (9).0009 Рис. 4а ).
Это наблюдение поддерживает эволюционную гипотезу о том, что Tn5053-подобный транспозон, содержащий TnsB, TnsC и TniQ, но не TnsA, ассимилировал и перепрофилировал эту систему CRISPR 19 . Следовательно, ожидается, что все CAST типа V-K будут действовать посредством репликативной транспозиции, приводя к существенной доле нежелательных продуктов коинтеграции. Таким образом, мы исследовали HELIX в качестве универсального подхода, позволяющего вставлять ДНК методом вырезания и вставки с другими V-K CAST различных типов (9).0009 Рис. 4а ). d . Схема AcHELIX с фланковой последовательностью ShCAST из 14 п.н. на pDonor. e , Покрытие продуктов вставки в геном (TS2) в результате секвенирования с длинным считыванием, отображающее выборку типичных простых считываний вставок для AcHELIX и чтений коинтеграции для AcCAST (покрытие из чтений коинтеграции AcHELIX и чтений простых вставок AcCAST опущено для простоты). Транспонированные продукты были обогащены перед секвенированием с помощью Cas9.целенаправленное обогащение. f , Распределение длины считывания продуктов транспонирования при использовании AcCAST и AcHELIX на TS2 из данных секвенирования длительного считывания. Верхняя правая панель представляет собой увеличенное представление пика длины считывания ~8,3 т.п.н. г , Сравнение пропорций продукта простой вставки и коинтеграции для AcCAST и AcHELIX, оцененное с помощью секвенирования с длительным чтением. h , i , Эффективность интеграции в ориентациях T-LR и T-RL (панели h и и соответственно) в шести геномных целевых сайтах для AcCAST и AcHELIX, оцененных с помощью ddPCR.
d . Схема AcHELIX с фланковой последовательностью ShCAST из 14 п.н. на pDonor. e , Покрытие продуктов вставки в геном (TS2) в результате секвенирования с длинным считыванием, отображающее выборку типичных простых считываний вставок для AcHELIX и чтений коинтеграции для AcCAST (покрытие из чтений коинтеграции AcHELIX и чтений простых вставок AcCAST опущено для простоты). Транспонированные продукты были обогащены перед секвенированием с помощью Cas9.целенаправленное обогащение. f , Распределение длины считывания продуктов транспонирования при использовании AcCAST и AcHELIX на TS2 из данных секвенирования длительного считывания. Верхняя правая панель представляет собой увеличенное представление пика длины считывания ~8,3 т.п.н. г , Сравнение пропорций продукта простой вставки и коинтеграции для AcCAST и AcHELIX, оцененное с помощью секвенирования с длительным чтением. h , i , Эффективность интеграции в ориентациях T-LR и T-RL (панели h и и соответственно) в шести геномных целевых сайтах для AcCAST и AcHELIX, оцененных с помощью ddPCR. На панели h эффективность интеграции AcHELIX T-LR по сравнению с AcCAST показана на правой панели. Все трансформации содержат вариант pDonor с флангами ShCAST и интервалом 14 п.н. между сайтами nAniI и LE/RE. j , Эффективность интеграции при использовании AcHELIX с использованием sgRNA, нацеленной на TS6, и pDonors, кодирующих грузы различных размеров, оценивали с помощью ddPCR. к , Схема ShoHELIX с фланковой последовательностью ShCAST из 14 п.н. на pDonor. l , Охват ожидаемых продуктов вставки в геном (TS2) в результате секвенирования с длительным считыванием, отображающий выборку образцов простых считываний вставок для ShoHELIX и коинтегрированных чтений для ShoCAST (покрытие из коинтегрированных считываний ShoHELIX и простых вставок ShoCAST чтения опущены для простоты). Транспонированные продукты были обогащены перед секвенированием с помощью обогащения мишени Cas9. m , Распределение длины чтения при использовании ShoCAST и ShoHELIX на геномной мишени (TS2) из данных секвенирования длительного чтения.
На панели h эффективность интеграции AcHELIX T-LR по сравнению с AcCAST показана на правой панели. Все трансформации содержат вариант pDonor с флангами ShCAST и интервалом 14 п.н. между сайтами nAniI и LE/RE. j , Эффективность интеграции при использовании AcHELIX с использованием sgRNA, нацеленной на TS6, и pDonors, кодирующих грузы различных размеров, оценивали с помощью ddPCR. к , Схема ShoHELIX с фланковой последовательностью ShCAST из 14 п.н. на pDonor. l , Охват ожидаемых продуктов вставки в геном (TS2) в результате секвенирования с длительным считыванием, отображающий выборку образцов простых считываний вставок для ShoHELIX и коинтегрированных чтений для ShoCAST (покрытие из коинтегрированных считываний ShoHELIX и простых вставок ShoCAST чтения опущены для простоты). Транспонированные продукты были обогащены перед секвенированием с помощью обогащения мишени Cas9. m , Распределение длины чтения при использовании ShoCAST и ShoHELIX на геномной мишени (TS2) из данных секвенирования длительного чтения. n , Сравнение пропорций продуктов для простой вставки и коинтеграции для ShoCAST и ShoHELIX, оцененных с помощью секвенирования с длительным чтением. o , p , Эффективность интеграции в ориентациях T-LR и T-RL (панели o и p соответственно) в шести геномных целевых сайтах для ShoCAST и ShoHELIX, оцененная с помощью ddPCR. q , Эффективность интеграции при использовании ShoHELIX с sgRNA, нацеленной на TS3, и pDonors, кодирующих грузы различных размеров, оценивалась с помощью ddPCR. Все трансформации ShoCAST и ShoHELIX содержат вариант pDonor с флангами ShCAST. Для 9Панели 0009 c, h — j и o — q , среднее значение, стандартное отклонение и отдельные точки данных показаны для n = 3. LE и RE, левый и правый концы транспозона соответственно; sgRNA, одиночная направляющая РНК.
n , Сравнение пропорций продуктов для простой вставки и коинтеграции для ShoCAST и ShoHELIX, оцененных с помощью секвенирования с длительным чтением. o , p , Эффективность интеграции в ориентациях T-LR и T-RL (панели o и p соответственно) в шести геномных целевых сайтах для ShoCAST и ShoHELIX, оцененная с помощью ddPCR. q , Эффективность интеграции при использовании ShoHELIX с sgRNA, нацеленной на TS3, и pDonors, кодирующих грузы различных размеров, оценивалась с помощью ddPCR. Все трансформации ShoCAST и ShoHELIX содержат вариант pDonor с флангами ShCAST. Для 9Панели 0009 c, h — j и o — q , среднее значение, стандартное отклонение и отдельные точки данных показаны для n = 3. LE и RE, левый и правый концы транспозона соответственно; sgRNA, одиночная направляющая РНК. Во-первых, для канонической системы AcCAST мы разработали два каркаса sgRNA (9).0009 Fig. 4b ) и две архитектуры pDonor, последняя из которых отличалась тем, что содержала различные последовательности из 25 п.н., фланкирующие LE и RE (либо как сообщалось ранее для AcCAST 14 , либо с использованием фланкирующих последовательностей ShCAST). С двумя конструкциями sgRNA, которые различались в зависимости от их точек слияния crRNA-tracrRNA, мы наблюдали лишь скромную разницу в эффективности интеграции (, рис. 4b, и , 4c, ). Однако pDonor, содержащий фланкирующие последовательности ShCAST, приводил к повышенной абсолютной эффективности интеграции 190,6% или 20,4% для sgRNA-1 и sgRNA-2 соответственно (увеличение в 1,28 и 1,31 раза по сравнению с pDonor с нативными флангами AcCAST; , рис. 4c ). Как мы ранее наблюдали для ShCAST ( Sup. Fig. 1b ), эти результаты предполагают, что последовательности, непосредственно прилегающие к LE и RE на pDonor, являются важной детерминантой эффективности CAST-опосредованной интеграции типа V-K.
Во-первых, для канонической системы AcCAST мы разработали два каркаса sgRNA (9).0009 Fig. 4b ) и две архитектуры pDonor, последняя из которых отличалась тем, что содержала различные последовательности из 25 п.н., фланкирующие LE и RE (либо как сообщалось ранее для AcCAST 14 , либо с использованием фланкирующих последовательностей ShCAST). С двумя конструкциями sgRNA, которые различались в зависимости от их точек слияния crRNA-tracrRNA, мы наблюдали лишь скромную разницу в эффективности интеграции (, рис. 4b, и , 4c, ). Однако pDonor, содержащий фланкирующие последовательности ShCAST, приводил к повышенной абсолютной эффективности интеграции 190,6% или 20,4% для sgRNA-1 и sgRNA-2 соответственно (увеличение в 1,28 и 1,31 раза по сравнению с pDonor с нативными флангами AcCAST; , рис. 4c ). Как мы ранее наблюдали для ShCAST ( Sup. Fig. 1b ), эти результаты предполагают, что последовательности, непосредственно прилегающие к LE и RE на pDonor, являются важной детерминантой эффективности CAST-опосредованной интеграции типа V-K. Принимая во внимание последовательности флангов, а также другие потенциальные детерминанты эффективности интеграции, такие как число копий pDonor ( доп. Рис. 8 ), — это возможности для оптимизации различных CAST. Кроме того, AcCAST показал минимальное, хотя и все еще обнаруживаемое, количество вставок, ориентированных на T-RL, что делает его почти полным однонаправленным вставочным устройством (, рис. 4b, ).
Принимая во внимание последовательности флангов, а также другие потенциальные детерминанты эффективности интеграции, такие как число копий pDonor ( доп. Рис. 8 ), — это возможности для оптимизации различных CAST. Кроме того, AcCAST показал минимальное, хотя и все еще обнаруживаемое, количество вставок, ориентированных на T-RL, что делает его почти полным однонаправленным вставочным устройством (, рис. 4b, ). 4f, и , 4g, ). В шести геномных мишенях AcHELIX сохранил сравнимую с AcCAST интеграцию ДНК под управлением РНК и направленность вставки (, рис. 4h, и , 4i, ). Кроме того, как и ShHELIX, AcHELIX не продемонстрировал снижения эффективности при интеграции грузовых последовательностей различных размеров до 90,8 кб, сохраняя эффективность интеграции более 83% для всех четырех размеров груза в TS6 ( рис. 4j ). Таким образом, подобно ShHELIX, AcHELIX представляет собой эффективный сконструированный CAST с почти идеальной чистотой продукта простой вставки для вставок ДНК различных размеров.
4f, и , 4g, ). В шести геномных мишенях AcHELIX сохранил сравнимую с AcCAST интеграцию ДНК под управлением РНК и направленность вставки (, рис. 4h, и , 4i, ). Кроме того, как и ShHELIX, AcHELIX не продемонстрировал снижения эффективности при интеграции грузовых последовательностей различных размеров до 90,8 кб, сохраняя эффективность интеграции более 83% для всех четырех размеров груза в TS6 ( рис. 4j ). Таким образом, подобно ShHELIX, AcHELIX представляет собой эффективный сконструированный CAST с почти идеальной чистотой продукта простой вставки для вставок ДНК различных размеров. Характеристика продуктов вставки с помощью секвенирования с длительным чтением выявила 54,61% коинтегратов для ShoCAST и 20,54% для ShoHELIX, демонстрируя 2,6-кратное снижение коинтегратов при использовании ShoHELIX (, рис. 4l-4m ). Для геномных мишеней TS2-TS7 мы наблюдали диапазон эффективности интеграции, при этом ShoHELIX демонстрировал сопоставимую интеграцию с ShoCAST (, рис. 4o, ). Подобно AcCAST и AcHELIX, направленность вставок ShoCAST и ShoHELIX была преимущественно в ориентации T-LR, хотя и с минимальными, но обнаруживаемыми вставками T-RL (9).0009 Рис. 4o и 4p ). Кроме того, в отличие от ShHELIX и AcHELIX, ShoHELIX показал снижение эффективности интеграции с увеличением размера груза на pDonor при TS3 (, рис. 4q, ).
Характеристика продуктов вставки с помощью секвенирования с длительным чтением выявила 54,61% коинтегратов для ShoCAST и 20,54% для ShoHELIX, демонстрируя 2,6-кратное снижение коинтегратов при использовании ShoHELIX (, рис. 4l-4m ). Для геномных мишеней TS2-TS7 мы наблюдали диапазон эффективности интеграции, при этом ShoHELIX демонстрировал сопоставимую интеграцию с ShoCAST (, рис. 4o, ). Подобно AcCAST и AcHELIX, направленность вставок ShoCAST и ShoHELIX была преимущественно в ориентации T-LR, хотя и с минимальными, но обнаруживаемыми вставками T-RL (9).0009 Рис. 4o и 4p ). Кроме того, в отличие от ShHELIX и AcHELIX, ShoHELIX показал снижение эффективности интеграции с увеличением размера груза на pDonor при TS3 (, рис. 4q, ). Однако открытые и охарактеризованные в настоящее время системы типа I и В-К имеют ограничения, ограничивающие простоту их использования, в том числе размер ( доп. рис. 9 ), стехиометрическая сложность и сложность компонентов и/или чистота продукта введения. Чтобы преодолеть эти ограничения, мы разработали HELIX, которая использует технологические преимущества CAST типа V-K и использует слияние nHE и модифицированную донорную плазмиду для достижения программируемой и эффективной вставки ДНК путем вырезания и вставки, аналогичной CAST типа I. Мы показываем, что HELIX резко повышает чистоту продукта простой вставки на плазмидных и геномных мишенях в E. coli и сохраняет надежную транспозицию под управлением РНК на уровне дикого типа или близком к нему. Кроме того, мы показываем возможность упрощения систем CAST и HELIX до трехкомпонентных систем путем слияния субблоков с Cas12k, полезность которых будет улучшена будущими инженерными усилиями по повышению эффективности их интеграции.
Однако открытые и охарактеризованные в настоящее время системы типа I и В-К имеют ограничения, ограничивающие простоту их использования, в том числе размер ( доп. рис. 9 ), стехиометрическая сложность и сложность компонентов и/или чистота продукта введения. Чтобы преодолеть эти ограничения, мы разработали HELIX, которая использует технологические преимущества CAST типа V-K и использует слияние nHE и модифицированную донорную плазмиду для достижения программируемой и эффективной вставки ДНК путем вырезания и вставки, аналогичной CAST типа I. Мы показываем, что HELIX резко повышает чистоту продукта простой вставки на плазмидных и геномных мишенях в E. coli и сохраняет надежную транспозицию под управлением РНК на уровне дикого типа или близком к нему. Кроме того, мы показываем возможность упрощения систем CAST и HELIX до трехкомпонентных систем путем слияния субблоков с Cas12k, полезность которых будет улучшена будущими инженерными усилиями по повышению эффективности их интеграции. Вместе наши подходы являются первым описанием технологии CAST и подчеркивают, как можно использовать другие встречающиеся в природе ферменты для улучшения свойств CAST.
Вместе наши подходы являются первым описанием технологии CAST и подчеркивают, как можно использовать другие встречающиеся в природе ферменты для улучшения свойств CAST.
 Параллельно с этим HELIX обладает потенциалом для расширения использования CAST для различных организмов, учитывая его улучшенную субъединицу, стехиометрию и простоту механизма вставки. В целом, HELIX демонстрирует перспективу применения инженерных подходов для оптимизации и преодоления ограничений CAST с последствиями для разработки оптимальных технологий вставки ДНК.
Параллельно с этим HELIX обладает потенциалом для расширения использования CAST для различных организмов, учитывая его улучшенную субъединицу, стехиометрию и простоту механизма вставки. В целом, HELIX демонстрирует перспективу применения инженерных подходов для оптимизации и преодоления ограничений CAST с последствиями для разработки оптимальных технологий вставки ДНК.
 Фрагменты гена для клонирования конструкции были заказаны в Twist Biosciences; синтетический SpCas9sgRNA были заказаны в Synthego (, дополнительная таблица 2, ).
Фрагменты гена для клонирования конструкции были заказаны в Twist Biosciences; синтетический SpCas9sgRNA были заказаны в Synthego (, дополнительная таблица 2, ). 5.0.16; Qiagen).
5.0.16; Qiagen). События вставки измеряли с использованием целевых праймеров и донор-специфического зонда (, дополнительная таблица 3, ). Реакции ddPCR содержали 20 пг плазмидной ДНК или 2 нг гДНК, 250 нМ каждого праймера, 900 нМ зонда и супермикс ddPCR для зондов (без dUTP) (BioRad) в 20 мкл реакций, а капли генерировали с использованием автоматического генератора капель QX200 ( БиоРад). Условия термоциклирования: 1 цикл (95°С 10 мин), 40 циклов (94°С 30 сек, 58°С 1 мин), 1 цикл (98°С 10 мин), выдержка при 4°С. Продукты ПЦР анализировали с помощью устройства для считывания капель QX200 (BioRad), а абсолютное количество вставок определяли с помощью QuantaSoft (v1.7.4). Также анализировали общую матричную ДНК и рассчитывали эффективность интеграции по вставкам/матрице*100.
События вставки измеряли с использованием целевых праймеров и донор-специфического зонда (, дополнительная таблица 3, ). Реакции ddPCR содержали 20 пг плазмидной ДНК или 2 нг гДНК, 250 нМ каждого праймера, 900 нМ зонда и супермикс ddPCR для зондов (без dUTP) (BioRad) в 20 мкл реакций, а капли генерировали с использованием автоматического генератора капель QX200 ( БиоРад). Условия термоциклирования: 1 цикл (95°С 10 мин), 40 циклов (94°С 30 сек, 58°С 1 мин), 1 цикл (98°С 10 мин), выдержка при 4°С. Продукты ПЦР анализировали с помощью устройства для считывания капель QX200 (BioRad), а абсолютное количество вставок определяли с помощью QuantaSoft (v1.7.4). Также анализировали общую матричную ДНК и рассчитывали эффективность интеграции по вставкам/матрице*100. Транспонированные продукты обогащали путем электропорации примерно 100 нг плазмидного пула в электрокомпетентные клетки Endura (Lucigen), которые представляют собой не-PIR штамм, который ограничивает рекомбинацию. Клетки восстанавливали в течение 1 часа при 37°C в S.O.C. и распределяли по чашкам с агаром LB, содержащим 50 мкг/мл канамицина и 25 мкг/мл хлорамфеникола. Планшеты инкубировали при 30°C (для ограничения рекомбинации) в течение 24 часов, соскребали и экстрагировали плазмидную ДНК с помощью минипрепарата. Обогащенные плазмиды расщепляли EcoRV (NEB) в течение 8 часов при 37°С. Подготовка библиотеки для секвенирования без амплификации (Oxford Nanopore Technologies, SQK-LSK109)) выполняли с использованием набора расширения штрих-кода (Oxford Nanopore Technologies, NBD-104). Окончательную объединенную библиотеку загружали в проточную кювету R9.4.1 и секвенировали в течение 24 часов.
Транспонированные продукты обогащали путем электропорации примерно 100 нг плазмидного пула в электрокомпетентные клетки Endura (Lucigen), которые представляют собой не-PIR штамм, который ограничивает рекомбинацию. Клетки восстанавливали в течение 1 часа при 37°C в S.O.C. и распределяли по чашкам с агаром LB, содержащим 50 мкг/мл канамицина и 25 мкг/мл хлорамфеникола. Планшеты инкубировали при 30°C (для ограничения рекомбинации) в течение 24 часов, соскребали и экстрагировали плазмидную ДНК с помощью минипрепарата. Обогащенные плазмиды расщепляли EcoRV (NEB) в течение 8 часов при 37°С. Подготовка библиотеки для секвенирования без амплификации (Oxford Nanopore Technologies, SQK-LSK109)) выполняли с использованием набора расширения штрих-кода (Oxford Nanopore Technologies, NBD-104). Окончательную объединенную библиотеку загружали в проточную кювету R9.4.1 и секвенировали в течение 24 часов. 2 ). Полученная библиотека была загружена на R9..4.1 проточная ячейка и секвенирование в течение 30 часов.
2 ). Полученная библиотека была загружена на R9..4.1 проточная ячейка и секвенирование в течение 30 часов. Графики покрытия были созданы на основе примерного набора из 100 прочтений с использованием Geneious (v2021.2.2) и его встроенного выравнивателя (средняя чувствительность и повторение до 5 раз). Также были созданы файлы Sam, содержащие выровненные чтения, которые использовались для создания гистограмм длины.
Графики покрытия были созданы на основе примерного набора из 100 прочтений с использованием Geneious (v2021.2.2) и его встроенного выравнивателя (средняя чувствительность и повторение до 5 раз). Также были созданы файлы Sam, содержащие выровненные чтения, которые использовались для создания гистограмм длины. Продукты ПЦР перед количественным определением очищали с помощью QuantiFluor (Promega) и объединяли в эквимолярный пул. Окончательные библиотеки количественно оценивали с помощью количественной ПЦР (набор для количественной оценки KAPA Library; Roche 7960140001) и секвенировали на MiSeq с использованием набора v2 на 300 циклов (Illumina).
Продукты ПЦР перед количественным определением очищали с помощью QuantiFluor (Promega) и объединяли в эквимолярный пул. Окончательные библиотеки количественно оценивали с помощью количественной ПЦР (набор для количественной оценки KAPA Library; Roche 7960140001) и секвенировали на MiSeq с использованием набора v2 на 300 циклов (Illumina). Т. Хилле, C.R.R. Алвес и Р.А. Silverstein за предложения по рукописи, P.M. Boone за помощь с ddPCR и B.L. Стоддард за советом. C.J.T. был поддержан грантом № 2020295403 Национального научного фонда для аспирантов. BPK был поддержан стипендией Говарда М. Гудмана Mass General Hospital.
Т. Хилле, C.R.R. Алвес и Р.А. Silverstein за предложения по рукописи, P.M. Boone за помощь с ddPCR и B.L. Стоддард за советом. C.J.T. был поддержан грантом № 2020295403 Национального научного фонда для аспирантов. BPK был поддержан стипендией Говарда М. Гудмана Mass General Hospital. Нац. Ред. Микробиол . 11, 435–442 (2013).
Нац. Ред. Микробиол . 11, 435–442 (2013).  and Jasin, M. Экспрессия сайт-специфичной эндонуклеазы стимулирует гомологичную рекомбинацию в клетках млекопитающих. Труды Национальной академии наук 91, 6064–6068 (1994).
and Jasin, M. Экспрессия сайт-специфичной эндонуклеазы стимулирует гомологичную рекомбинацию в клетках млекопитающих. Труды Национальной академии наук 91, 6064–6068 (1994). 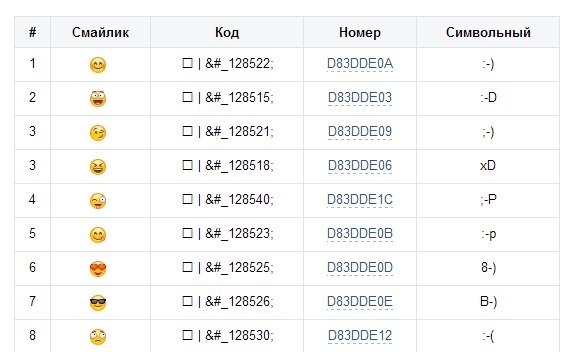 Природа 571, 219–225 (2019).
Природа 571, 219–225 (2019). 
 Природа (2021).
Природа (2021).  Труды Национальной академии наук 110, (2013).
Труды Национальной академии наук 110, (2013). 

 Даже в холмистом Окленде вы не найдете чистых подъемов на высоту 1000 метров.
Даже в холмистом Окленде вы не найдете чистых подъемов на высоту 1000 метров. ч) + 1 мин RI @ легкий (уклон 3–5 % на 8 км.ч))
ч) + 1 мин RI @ легкий (уклон 3–5 % на 8 км.ч))  Обратите внимание, как 30 секунд задают скорость.
Обратите внимание, как 30 секунд задают скорость.  ч + 2 мин. RI (6% 7 км.ч)
ч + 2 мин. RI (6% 7 км.ч)  ) Легкое регулирование, но тяжелая фильтрация — это слово дня.
) Легкое регулирование, но тяжелая фильтрация — это слово дня. Скорее, Виктор использует «интеллектуальный» подход к контролю вибрации. Например, нижнее шасси соединено с вибропоглощающей пластиной ½ дюйма, которая является неотъемлемой частью конструкции. Плата ПК прикручена к этой пластине болтами в 17 местах!1328 против узлов. Как только они стали известны, чувствительные компоненты, такие как трубки и герметизированные конденсаторы типа «бумага в масле», были размещены в пучностях.
Скорее, Виктор использует «интеллектуальный» подход к контролю вибрации. Например, нижнее шасси соединено с вибропоглощающей пластиной ½ дюйма, которая является неотъемлемой частью конструкции. Плата ПК прикручена к этой пластине болтами в 17 местах!1328 против узлов. Как только они стали известны, чувствительные компоненты, такие как трубки и герметизированные конденсаторы типа «бумага в масле», были размещены в пучностях. Нагрузочный резистор 47 кОм постоянно подключен к входу картриджа по умолчанию. Предусмотрены гнездовые штыри, а также штырьковые выводы для вашего любимого аудиофильского колпачка или резистора. Фабрика будет рада помочь вам в этих вопросах.
Нагрузочный резистор 47 кОм постоянно подключен к входу картриджа по умолчанию. Предусмотрены гнездовые штыри, а также штырьковые выводы для вашего любимого аудиофильского колпачка или резистора. Фабрика будет рада помочь вам в этих вопросах. Я также попробовал несколько резисторов Holco разных номиналов, которые Виктор оставил мне. Мой вывод состоит в том, что нет жестких правил и , когда дело доходит до загрузки картриджа. Попробуйте это с вашим картриджем и послушайте результат. Поверьте мне, вы легко услышите разницу. Мой совет найти то, что тебе нравится и так далее. Не читайте ни меня, ни кого-либо другого, кто диктует вам, что вы должны делать в каждом случае. Там есть нет «на всякий случай».
Я также попробовал несколько резисторов Holco разных номиналов, которые Виктор оставил мне. Мой вывод состоит в том, что нет жестких правил и , когда дело доходит до загрузки картриджа. Попробуйте это с вашим картриджем и послушайте результат. Поверьте мне, вы легко услышите разницу. Мой совет найти то, что тебе нравится и так далее. Не читайте ни меня, ни кого-либо другого, кто диктует вам, что вы должны делать в каждом случае. Там есть нет «на всякий случай». (Можем ли мы найти менее клинический способ описать это, интересно?) Да, это правда, аналог — это заноза в заднице, он занимает много времени, и нужно быть полностью одержимым и сумасшедшим, чтобы уйти. через все это. Но когда все в порядке… ах, чудо.
(Можем ли мы найти менее клинический способ описать это, интересно?) Да, это правда, аналог — это заноза в заднице, он занимает много времени, и нужно быть полностью одержимым и сумасшедшим, чтобы уйти. через все это. Но когда все в порядке… ах, чудо. 10.1182/blood-2008-09-179754 [PubMed] [CrossRef] [Google Scholar]
10.1182/blood-2008-09-179754 [PubMed] [CrossRef] [Google Scholar]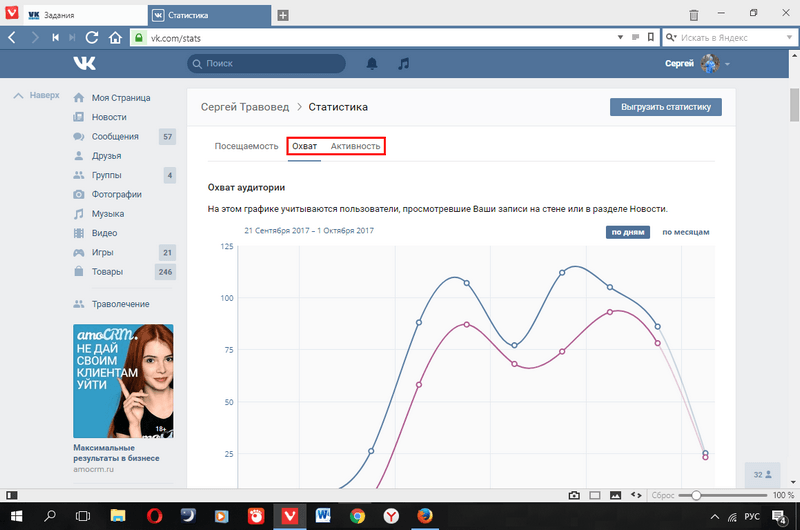 Crit Rev Immunol (2010) 30 (1): 1–29. 10.1615/CritRevImmunol.v30.i1.10 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Crit Rev Immunol (2010) 30 (1): 1–29. 10.1615/CritRevImmunol.v30.i1.10 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Clin Exp Immunol (1990) 81(3):357–67. 10.1111/j.1365-2249.1990.tb05339.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Clin Exp Immunol (1990) 81(3):357–67. 10.1111/j.1365-2249.1990.tb05339.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] , Смит К.И. Дефицит IgG2 у здорового донора крови. Сопутствующее отсутствие иммуноглобулинов IgG2, IgA и IgE и специфических антиуглеводных антител. Clin Exp Immunol (1983) 51(3):600–4. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Смит К.И. Дефицит IgG2 у здорового донора крови. Сопутствующее отсутствие иммуноглобулинов IgG2, IgA и IgE и специфических антиуглеводных антител. Clin Exp Immunol (1983) 51(3):600–4. [Бесплатная статья PMC] [PubMed] [Google Scholar] фон Гунтен С., Смит Д.Ф., Каммингс Р.Д., Ридель С., Мишер С., Шауб А. и соавт.
Внутривенный иммуноглобулин содержит широкий спектр антиуглеводных антител, который не ограничивается подклассом IgG2. J Allergy Clin Immunol (2009) 123(6):1268–76. 10.1016/j.jaci.2009.03.013 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
фон Гунтен С., Смит Д.Ф., Каммингс Р.Д., Ридель С., Мишер С., Шауб А. и соавт.
Внутривенный иммуноглобулин содержит широкий спектр антиуглеводных антител, который не ограничивается подклассом IgG2. J Allergy Clin Immunol (2009) 123(6):1268–76. 10.1016/j.jaci.2009.03.013 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Seppala IJ, Sarvas H, Makela O. Низкие концентрации аллотипических подмножеств Gm G3 mg и G1 mf у гомозигот и гетерозигот. Джей Иммунол (1993) 151(5):2529–37. [PubMed] [Google Scholar]
Seppala IJ, Sarvas H, Makela O. Низкие концентрации аллотипических подмножеств Gm G3 mg и G1 mf у гомозигот и гетерозигот. Джей Иммунол (1993) 151(5):2529–37. [PubMed] [Google Scholar] Материнские антитела против антигенов группы крови плода A или B: литическая активность подклассов IgG при цитотоксичности, обусловленной моноцитами, и корреляция с гемолитической болезнью новорожденных по системе ABO. Бр Дж. Гематол (1988) 70(4):465–9. 10.1111/j.1365-2141.1988.tb02518.x [PubMed] [CrossRef] [Google Scholar]
Материнские антитела против антигенов группы крови плода A или B: литическая активность подклассов IgG при цитотоксичности, обусловленной моноцитами, и корреляция с гемолитической болезнью новорожденных по системе ABO. Бр Дж. Гематол (1988) 70(4):465–9. 10.1111/j.1365-2141.1988.tb02518.x [PubMed] [CrossRef] [Google Scholar] Pediatr Allergy Immunol (2006) 17(7):544–50. 10.1111/j.1399-3038.2006.00454.x [PubMed] [CrossRef] [Google Scholar]
Pediatr Allergy Immunol (2006) 17(7):544–50. 10.1111/j.1399-3038.2006.00454.x [PubMed] [CrossRef] [Google Scholar] van Helden PM, van den Berg HM, Gouw SC, Kaijen PH, Zuurveld MG, Mauser-Bunschoten EP и др. .
Подклассы IgG антител против FVIII во время индукции иммунной толерантности у пациентов с гемофилией A. Br J Haematol (2008) 142(4):644–52. 10.1111/j.1365-2141.2008.07232.x [PubMed] [CrossRef] [Google Scholar]
van Helden PM, van den Berg HM, Gouw SC, Kaijen PH, Zuurveld MG, Mauser-Bunschoten EP и др. .
Подклассы IgG антител против FVIII во время индукции иммунной толерантности у пациентов с гемофилией A. Br J Haematol (2008) 142(4):644–52. 10.1111/j.1365-2141.2008.07232.x [PubMed] [CrossRef] [Google Scholar] 10.1007/s10875-012-9705-0 [PubMed] [CrossRef] [Google Scholar]
10.1007/s10875-012-9705-0 [PubMed] [CrossRef] [Google Scholar] Yamamoto M, Tabeya T, Naishiro Y, Yajima H, Ishigami K, Shimizu Y, et al.
Значение сывороточного IgG4 в диагностике заболеваний, связанных с IgG4, и в дифференциации от ревматических заболеваний и других заболеваний. Мод Ревматол (2012) 22 (3): 419–25. 10.1007/s10165-011-0532-6 [PubMed] [CrossRef] [Google Scholar]
Yamamoto M, Tabeya T, Naishiro Y, Yajima H, Ishigami K, Shimizu Y, et al.
Значение сывороточного IgG4 в диагностике заболеваний, связанных с IgG4, и в дифференциации от ревматических заболеваний и других заболеваний. Мод Ревматол (2012) 22 (3): 419–25. 10.1007/s10165-011-0532-6 [PubMed] [CrossRef] [Google Scholar] 10.1073/pnas.75.5.2429 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10.1073/pnas.75.5.2429 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Мол Иммунол (2002) 38(14):1073–83. 10.1016/S0161-5890(02)00036-6 [PubMed] [CrossRef] [Google Scholar]
Мол Иммунол (2002) 38(14):1073–83. 10.1016/S0161-5890(02)00036-6 [PubMed] [CrossRef] [Google Scholar] Proc Natl Acad Sci U S A (2008) 105(16):6045–50. 10.1073/пнас.0800159105 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Proc Natl Acad Sci U S A (2008) 105(16):6045–50. 10.1073/пнас.0800159105 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Естественные вариации гликозилирования Fc ВИЧ-специфических антител влияют на противовирусную активность. Дж. Клин Инвест (2013) 123 (5): 2183–92. 10.1172/JCI65708DS1 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Естественные вариации гликозилирования Fc ВИЧ-специфических антител влияют на противовирусную активность. Дж. Клин Инвест (2013) 123 (5): 2183–92. 10.1172/JCI65708DS1 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] J Biol Chem (2007) 282(10):7405–15. 10.1074/jbc.M6026
J Biol Chem (2007) 282(10):7405–15. 10.1074/jbc.M6026 Biophys Chem (2001) 93 (2–3): 181–96. 10.1016/S0301-4622(01)00220-4 [PubMed] [CrossRef] [Google Scholar]
Biophys Chem (2001) 93 (2–3): 181–96. 10.1016/S0301-4622(01)00220-4 [PubMed] [CrossRef] [Google Scholar] Мутации в человеческом антителе IgG2 образуют отдельные и гомогенные дисульфидные изомеры, но не влияют на гамма-рецептор Fc или связывание C1q. Protein Sci (2010) 19(4):753–62. 10.1002/pro.352 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Мутации в человеческом антителе IgG2 образуют отдельные и гомогенные дисульфидные изомеры, но не влияют на гамма-рецептор Fc или связывание C1q. Protein Sci (2010) 19(4):753–62. 10.1002/pro.352 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Мол Иммунол (1993) 30(1):105–8. 10.1016/0161-5890(93)-B [PubMed] [CrossRef] [Google Scholar]
Мол Иммунол (1993) 30(1):105–8. 10.1016/0161-5890(93)-B [PubMed] [CrossRef] [Google Scholar]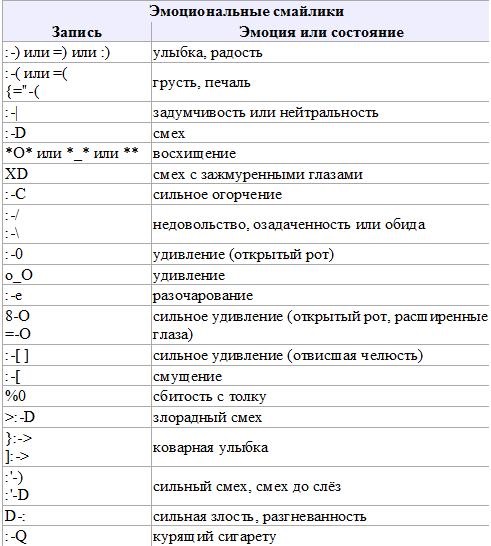 Дж. Биол. Химия (2014) 289(9): 6098–109. 10.1074/jbc.M113.541813 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Биол. Химия (2014) 289(9): 6098–109. 10.1074/jbc.M113.541813 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Г. Полиморфизмы иммуноглобулинов человека: аллотипы Gm, Am, Em и Km. Exp Clin Immunogenet (1989) 6(1):7–17. [PubMed] [Google Scholar]
Г. Полиморфизмы иммуноглобулинов человека: аллотипы Gm, Am, Em и Km. Exp Clin Immunogenet (1989) 6(1):7–17. [PubMed] [Google Scholar]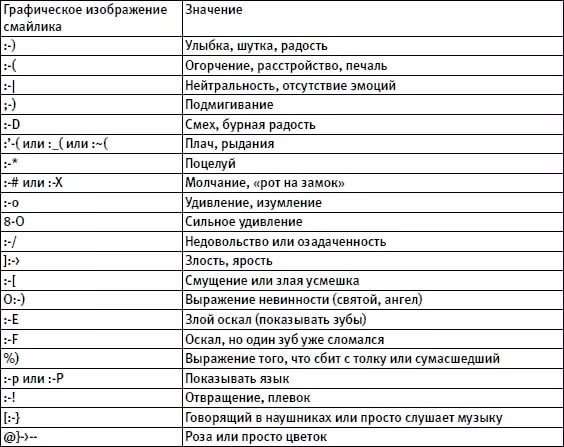 Иммуногенетика (1995) 42(5):414–7. 10.1007/BF00179404 [PubMed] [CrossRef] [Google Scholar]
Иммуногенетика (1995) 42(5):414–7. 10.1007/BF00179404 [PubMed] [CrossRef] [Google Scholar] 10.1016/j.molimm.2011.09.002 [PubMed] [CrossRef] [Google Scholar]
10.1016/j.molimm.2011.09.002 [PubMed] [CrossRef] [Google Scholar]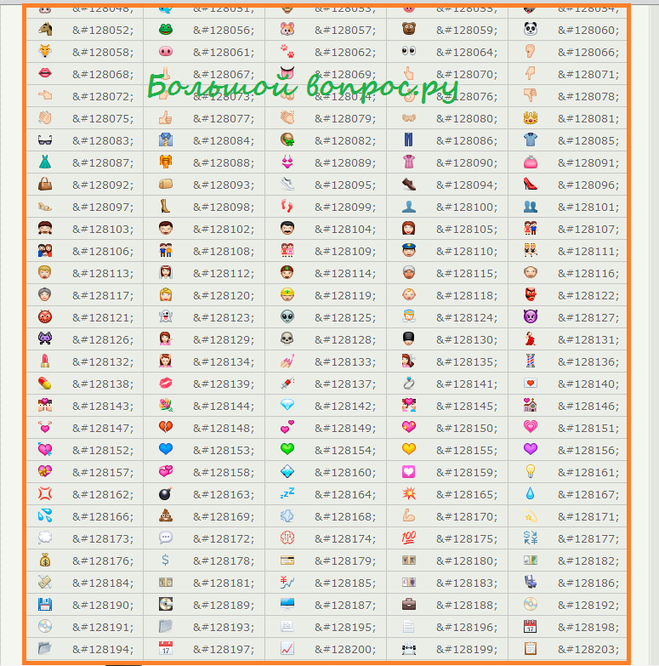 Arthritis Res Ther (2010) 12(6):R221. 10.1186/ar3208 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Arthritis Res Ther (2010) 12(6):R221. 10.1186/ar3208 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Nucleic Acids Res (2014) 42(8):4813–22. 10.1093/nar/gku159 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Nucleic Acids Res (2014) 42(8):4813–22. 10.1093/nar/gku159 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Parekh RB, Dwek RA, Sutton BJ, Fernandes DL, Leung A, Stanworth D, et al.
Ассоциация ревматоидного артрита и первичного остеоартрита с изменением паттерна гликозилирования общего сывороточного IgG. Природа (1985) 316(6027):452–7. 10.1038/316452a0 [PubMed] [CrossRef] [Google Scholar]
Parekh RB, Dwek RA, Sutton BJ, Fernandes DL, Leung A, Stanworth D, et al.
Ассоциация ревматоидного артрита и первичного остеоартрита с изменением паттерна гликозилирования общего сывороточного IgG. Природа (1985) 316(6027):452–7. 10.1038/316452a0 [PubMed] [CrossRef] [Google Scholar] Menni C, Keser T, Mangino M, Bell JT, Erte I, Akmacic I, et al.
Гликозилирование иммуноглобулинов: роль генетических и эпигенетических влияний. PLoS One (2013) 8(12):e82558. 10.1371/journal.pone.0082558 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Menni C, Keser T, Mangino M, Bell JT, Erte I, Akmacic I, et al.
Гликозилирование иммуноглобулинов: роль генетических и эпигенетических влияний. PLoS One (2013) 8(12):e82558. 10.1371/journal.pone.0082558 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Immunol Lett (2011) 137 (1–2): 28–37. 10.1016/j.imlet.2011.01.017 [PubMed] [CrossRef] [Google Scholar]
Immunol Lett (2011) 137 (1–2): 28–37. 10.1016/j.imlet.2011.01.017 [PubMed] [CrossRef] [Google Scholar] Genes Cells (2011) 16(11):1071–80. 10.1111/j.1365-2443.2011.01552.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Genes Cells (2011) 16(11):1071–80. 10.1111/j.1365-2443.2011.01552.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Опытный гематол (2009) 37 (3): 309–21. 10.1016/j.exphem.2008.11.006 [PubMed] [CrossRef] [Google Scholar]
Опытный гематол (2009) 37 (3): 309–21. 10.1016/j.exphem.2008.11.006 [PubMed] [CrossRef] [Google Scholar] Zou G, Ochiai H, Huang W, Yang Q, Li C, Wang LX. Хемоферментативный синтез и связывание с рецептором Fcgamma гомогенных гликоформ Fc-домена антитела. Присутствие разделенного пополам сахарного фрагмента усиливает сродство Fc к рецептору FcgammaIIIa. J Am Chem Soc (2011) 133 (46): 18975–91. 10.1021/ja208390n [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Zou G, Ochiai H, Huang W, Yang Q, Li C, Wang LX. Хемоферментативный синтез и связывание с рецептором Fcgamma гомогенных гликоформ Fc-домена антитела. Присутствие разделенного пополам сахарного фрагмента усиливает сродство Fc к рецептору FcgammaIIIa. J Am Chem Soc (2011) 133 (46): 18975–91. 10.1021/ja208390n [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 10.1002/bit.20777 [PubMed] [CrossRef] [Google Scholar]
10.1002/bit.20777 [PubMed] [CrossRef] [Google Scholar] Наука (2006) 313 (5787): 670–3. 10.1126/наука.1129594 [PubMed] [CrossRef] [Google Scholar]
Наука (2006) 313 (5787): 670–3. 10.1126/наука.1129594 [PubMed] [CrossRef] [Google Scholar] Биохимия (1976) 15 (23): 5175–81. 10.1021/bi00668a035 [PubMed] [CrossRef] [Google Scholar]
Биохимия (1976) 15 (23): 5175–81. 10.1021/bi00668a035 [PubMed] [CrossRef] [Google Scholar] Аминокислотные остатки, специфичные для изотипа IgG человека, влияющие на комплемент-опосредованный лизис клеток и фагоцитоз. Eur J Immunol (1994) 24(10):2542–7. 10.1002/eji.1830241042 [PubMed] [CrossRef] [Google Scholar]
Аминокислотные остатки, специфичные для изотипа IgG человека, влияющие на комплемент-опосредованный лизис клеток и фагоцитоз. Eur J Immunol (1994) 24(10):2542–7. 10.1002/eji.1830241042 [PubMed] [CrossRef] [Google Scholar] Биофиз J (2007) 93 (11): 3733–44. 10.1529/biophysj.107.108993 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Биофиз J (2007) 93 (11): 3733–44. 10.1529/biophysj.107.108993 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Biochem J (2010) 432(1):101–11. 10.1042/BJ20100641 [PubMed] [CrossRef] [Google Scholar]
Biochem J (2010) 432(1):101–11. 10.1042/BJ20100641 [PubMed] [CrossRef] [Google Scholar] J Immunol (2011) 187(6):3208–17. 10.4049/jimmunol.1101467 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
J Immunol (2011) 187(6):3208–17. 10.4049/jimmunol.1101467 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Мол Иммунол (1998) 35(14–15):989–96. 10.1016/S0161-5890(98)00069-8 [PubMed] [CrossRef] [Google Scholar]
Мол Иммунол (1998) 35(14–15):989–96. 10.1016/S0161-5890(98)00069-8 [PubMed] [CrossRef] [Google Scholar] J Biol Chem (2001) 276(9): 6591–604. 10.1074/jbc.M009483200 [PubMed] [CrossRef] [Google Scholar]
J Biol Chem (2001) 276(9): 6591–604. 10.1074/jbc.M009483200 [PubMed] [CrossRef] [Google Scholar] О взаимодействии подклассов IgG с низкоаффинным Fc гамма RIIa (CD32) на моноцитах, нейтрофилах и тромбоцитах человека. Анализ функционального полиморфизма IgG2 человека. J Clin Invest (1992) 90 (4): 1537–46. 10.1172/JCI116022 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
О взаимодействии подклассов IgG с низкоаффинным Fc гамма RIIa (CD32) на моноцитах, нейтрофилах и тромбоцитах человека. Анализ функционального полиморфизма IgG2 человека. J Clin Invest (1992) 90 (4): 1537–46. 10.1172/JCI116022 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Фенотипическая изменчивость рецепторов IgG неклассическими аллелями FCGR2C. J Immunol (2012) 188(3):1318–24. 10.4049/jimmunol.1003945 [PubMed] [CrossRef] [Google Scholar]
Фенотипическая изменчивость рецепторов IgG неклассическими аллелями FCGR2C. J Immunol (2012) 188(3):1318–24. 10.4049/jimmunol.1003945 [PubMed] [CrossRef] [Google Scholar] 10.1016/j.clim.2005.10.008 [PubMed] [CrossRef] [Google Scholar]
10.1016/j.clim.2005.10.008 [PubMed] [CrossRef] [Google Scholar] Аномально короткие периоды полураспада IgG в сыворотке у мышей с дефицитом бета-2-микроглобулина. Eur J Immunol (1996) 26(3):690–6. 10.1002/eji.1830260327 [PubMed] [CrossRef] [Google Scholar]
Аномально короткие периоды полураспада IgG в сыворотке у мышей с дефицитом бета-2-микроглобулина. Eur J Immunol (1996) 26(3):690–6. 10.1002/eji.1830260327 [PubMed] [CrossRef] [Google Scholar]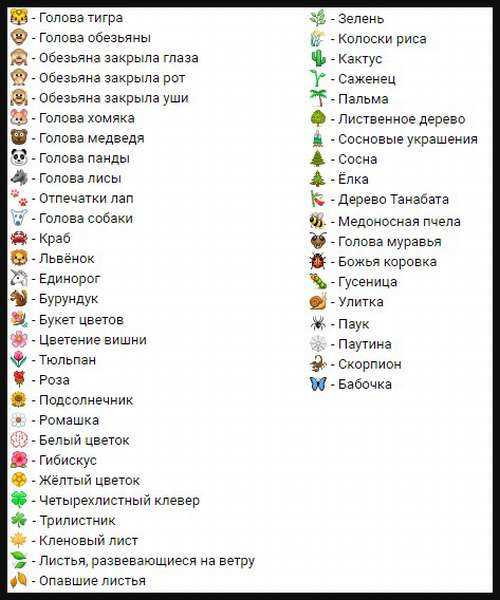 J Immunol (2003) 170(7):3528–33. 10.4049/jimmunol.170.7.3528 [PubMed] [CrossRef] [Google Scholar]
J Immunol (2003) 170(7):3528–33. 10.4049/jimmunol.170.7.3528 [PubMed] [CrossRef] [Google Scholar] 10.1111/trf.12334 [PubMed] [CrossRef] [Google Scholar]
10.1111/trf.12334 [PubMed] [CrossRef] [Google Scholar]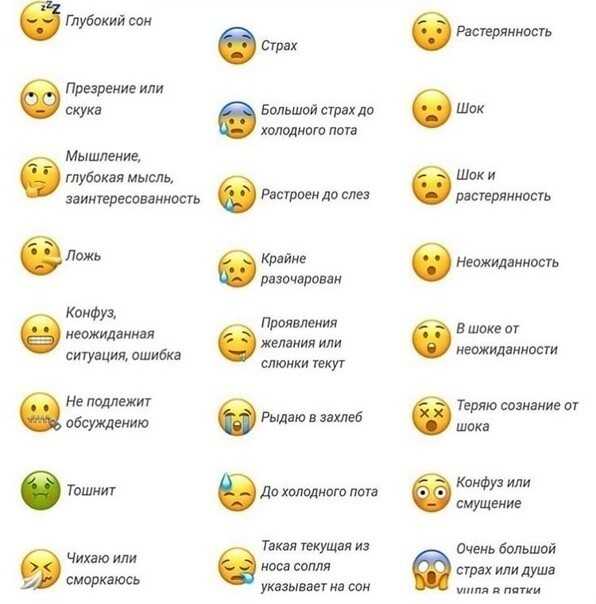 MAbs (2011) 3(5):431–9. 10.4161/mabs.3.5.17023 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
MAbs (2011) 3(5):431–9. 10.4161/mabs.3.5.17023 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Ладинский М.С., Хьюи-Тубман К.Е., Бьоркман П.Дж. Электронная томография поздних стадий FcRn-опосредованного трансцитоза антител в тонкой кишке новорожденных крыс. Mol Biol Cell (2012) 23(13):2537–45. 10.1091/mbc.E12-02-0093 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Ладинский М.С., Хьюи-Тубман К.Е., Бьоркман П.Дж. Электронная томография поздних стадий FcRn-опосредованного трансцитоза антител в тонкой кишке новорожденных крыс. Mol Biol Cell (2012) 23(13):2537–45. 10.1091/mbc.E12-02-0093 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Двунаправленный FcRn-зависимый транспорт IgG в поляризованной линии эпителиальных клеток кишечника человека. Джей Клин Инвест (1999) 104(7):903–11. 10.1172/JCI6968 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Двунаправленный FcRn-зависимый транспорт IgG в поляризованной линии эпителиальных клеток кишечника человека. Джей Клин Инвест (1999) 104(7):903–11. 10.1172/JCI6968 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Yoshida M, Kobayashi K, Kuo TT, Bry L, Glickman JN, Claypool SM, et al.
Неонатальный Fc-рецептор для IgG регулирует иммунный ответ слизистой оболочки на люминальные бактерии. J Clin Invest (2006) 116 (8): 2142. 10.1172/JCI27821.2142 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Yoshida M, Kobayashi K, Kuo TT, Bry L, Glickman JN, Claypool SM, et al.
Неонатальный Fc-рецептор для IgG регулирует иммунный ответ слизистой оболочки на люминальные бактерии. J Clin Invest (2006) 116 (8): 2142. 10.1172/JCI27821.2142 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]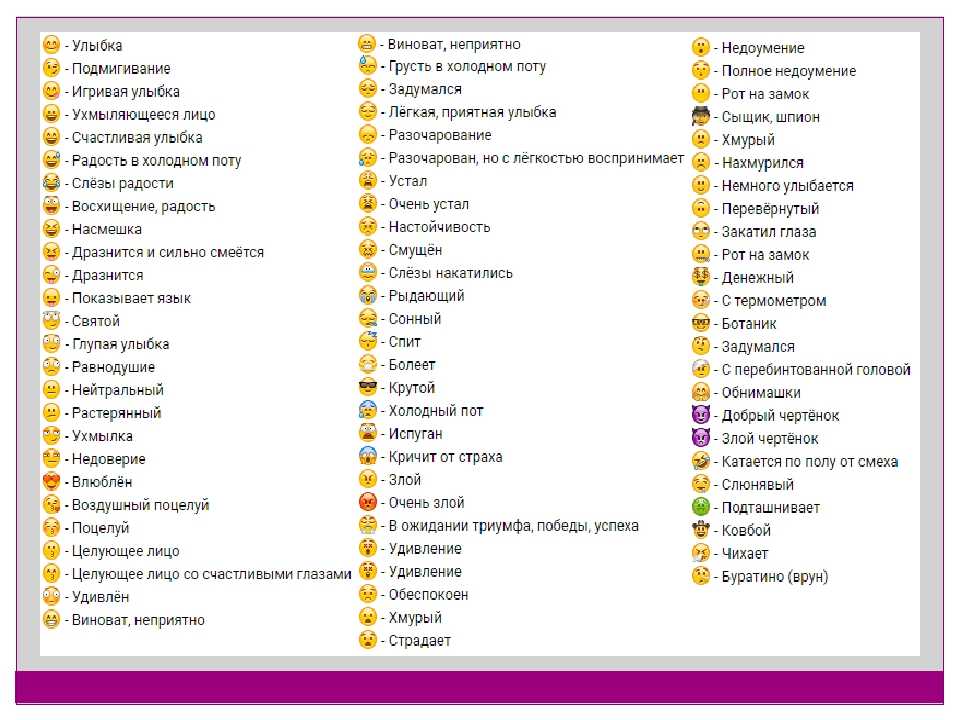 J Immunol (2011) 186(2):959–68. 10.4049/jimmunol.1000353 [PubMed] [CrossRef] [Google Scholar]
J Immunol (2011) 186(2):959–68. 10.4049/jimmunol.1000353 [PubMed] [CrossRef] [Google Scholar] Qiao SW, Kobayashi K, Johansen FE, Sollid LM, Andersen JT, Milford E, et al.
Зависимость опосредованной антителами презентации антигена от FcRn. Proc Natl Acad Sci U S A (2008) 105(27):9337–42. 10.1073/pnas.0801717105 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Qiao SW, Kobayashi K, Johansen FE, Sollid LM, Andersen JT, Milford E, et al.
Зависимость опосредованной антителами презентации антигена от FcRn. Proc Natl Acad Sci U S A (2008) 105(27):9337–42. 10.1073/pnas.0801717105 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Proc Natl Acad Sci USA (2007) 104(23):9770–5. 10.1073/pnas.0703354104 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Proc Natl Acad Sci USA (2007) 104(23):9770–5. 10.1073/pnas.0703354104 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Аутоантиген Ro52 напрямую взаимодействует с тяжелой цепью IgG человека in vivo в клетках млекопитающих. Мол Иммунол (2000) 37(10):591–602. 10.1016/S0161-5890(00)00068-7 [PubMed] [CrossRef] [Google Scholar]
Аутоантиген Ro52 напрямую взаимодействует с тяжелой цепью IgG человека in vivo в клетках млекопитающих. Мол Иммунол (2000) 37(10):591–602. 10.1016/S0161-5890(00)00068-7 [PubMed] [CrossRef] [Google Scholar] Rakebrandt N, Lentes S, Neumann H, James LC, Neumann-Staubitz P. Антитело- и TRIM21-зависимая внутриклеточная ограничение Salmonella enterica . Патог Дис (2014). 10.1111/2049-632X.12192 [PubMed] [CrossRef] [Google Scholar]
Rakebrandt N, Lentes S, Neumann H, James LC, Neumann-Staubitz P. Антитело- и TRIM21-зависимая внутриклеточная ограничение Salmonella enterica . Патог Дис (2014). 10.1111/2049-632X.12192 [PubMed] [CrossRef] [Google Scholar] 1038/ni.2548 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
1038/ni.2548 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Анализ молекулярного механизма терапии ВВИГ: взаимодействие между сывороточным IgG и DC-SIGN не зависит от гликоформы антитела или домена Fc. J Mol Biol (2013) 425 (8): 1253–8. 10.1016/j.jmb.2013.02.006 [PubMed] [CrossRef] [Google Scholar]
Анализ молекулярного механизма терапии ВВИГ: взаимодействие между сывороточным IgG и DC-SIGN не зависит от гликоформы антитела или домена Fc. J Mol Biol (2013) 425 (8): 1253–8. 10.1016/j.jmb.2013.02.006 [PubMed] [CrossRef] [Google Scholar]
