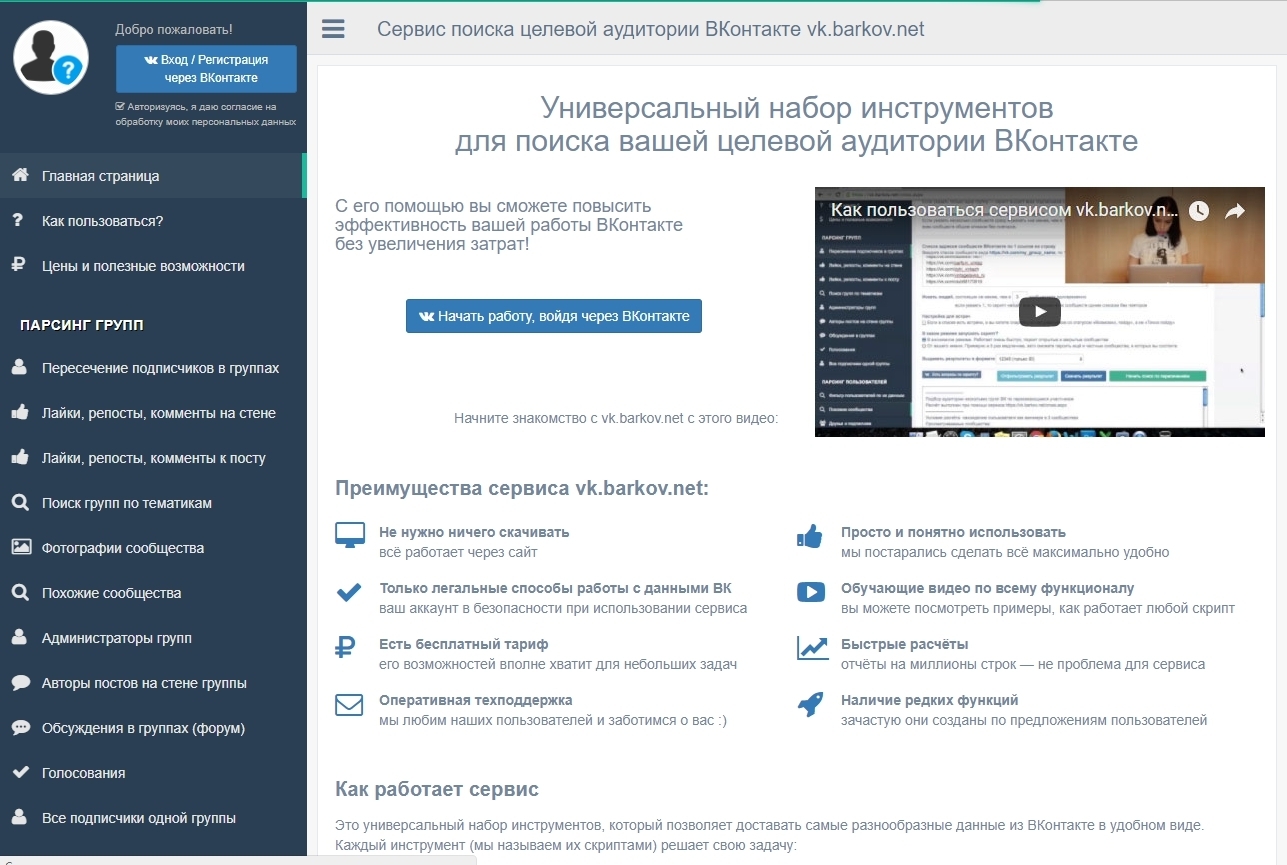
Преимущества VK Sova перед официальным софтом ВКонтакте
Хотя ВКонтакте предоставляет почти те же функции, что и Facebook, но, похоже, ему удается привлечь внимание людей. Согласно Alexa Ranking, VK занимает 15-е место, тогда как Facebook занимает 3-е место, а Twitter — 12-е место на момент, когда мы делаем этот обзор, то есть в феврале 2023 года.
Итак, да, теперь ВКонтакте стала третьей по посещаемости социальной сетью после Facebook и Twitter. Итак, да, это означает, что ВКонтакте теперь более популярен, чем Instagram, который занимает 16-е место в Alexa, LinkedIn — 30-е место и так далее.
Предыстория ВК
ВКонтакте базируется в Санкт-Петербурге, Россия. ВКонтакте также считается самой популярной социальной сетью не только в России, но и в Беларуси, Украине и Казахстане.
Основатель «Павел Дуров» запустил ВКонтакте 19 января 2007 года, и в феврале 2007 года ему удалось набрать 100 000 пользователей, которые выросли до 1 миллиона в июле 2007 года, а затем до 10 миллионов в апреле 2008 года. Да, это довольно быстро. К январю 2017 года у ВКонтакте было около 410 миллионов аккаунтов.
Да, это довольно быстро. К январю 2017 года у ВКонтакте было около 410 миллионов аккаунтов.
Особенности ВК
ВКонтакте позволяет пользователям общаться друг с другом публично или в частном порядке, создавать группы, общедоступные страницы и мероприятия, делиться и отмечать изображения, аудио и видео, играть в браузерные игры и так далее. ВКонтакте доступен на более чем 80 языках. ВКонтакте предоставляет мобильные приложения для Android, iPhone и WP.
На самом деле, ВКонтакте сделал свой сайт очень удобным. Те, кто использует сайты социальных сетей, такие как Facebook, могут очень легко использовать VK. Возможно, его простота делает его популярным, потому что люди всегда предпочитают удобные веб-сайты. Даже Условия использования ВКонтакте не такие строгие по сравнению с Facebook. ВКонтакте даже позволяет вам делиться контентом для взрослых. Так что, возможно, свобода — это еще одна точка притяжения к ВКонтакте. Ведь многим не нравятся строгие правила Facebook, а вам?
Да, блокировка контента для взрослых — это почти нормально, но Facebook все очень усложняет, например, у вас может быть только 5000 друзей, контент с большим количеством текста будет показан очень меньшему количеству друзей, контент со ссылкой на веб-сайт почти не виден ни в чьей ленте новостей если это не ваши близкие друзья, которые всегда продолжают болтать или комментировать ваш пост на Facebook, ваш пост на вашей фан-странице даже не отображается всем вашим подписчикам — только очень немногим подписчикам и так далее. Так что, возможно, эти ограничения Facebook привлекают пользователей социальных сетей к другим платформам, таким как VK, который почти такой же, как Facebook, но с гораздо меньшим ограничением вашей свободы.
Так что, возможно, эти ограничения Facebook привлекают пользователей социальных сетей к другим платформам, таким как VK, который почти такой же, как Facebook, но с гораздо меньшим ограничением вашей свободы.
Софт VK Sova
Для смартфонов на базе операционной системы Андроид было разработано специальное приложение «ВК Сова» — это неофициальная версия существующего ПО «ВКонтакте». Скачать ВК Сова бесплатно можно по ссылке.
Разработчики предлагают пользователям много интересных решений, отсутствующих в официальном клиенте. Легко заметить, что доработанный клиент почти ничем не отличается от официальной версии ПО для ВКонтакте. Главное отличие – это то, что можно делать загрузку музыки сразу на смартфон, чего нет в официальном приложении.
Интерфейс выглядит также, но есть отличия, просто они не видны на первый взгляд. Для получения доступа к дополнительным сервисам, найдите в настройках пункт «Сова». Там находятся все нестандартные функции для ВК.
При открытии этих опций можно включать доп. режимы пользования, либо отключать не нужные блоки, к примеру рекламу, можно применить нестандартные темы и многое другое.
режимы пользования, либо отключать не нужные блоки, к примеру рекламу, можно применить нестандартные темы и многое другое.
Где найти toggles в VK для Android и как их включать – Telegraph
Telegram-канал VK Leaks • t.me/vk_leaksToggles — это переключатели, с помощью которых можно управлять множеством функций, имеющихся в приложении ВКонтакте для Android.
Там могут прятаться как пока еще скрытые не вышедшие новинки, так и полезные опции.
И так, где найти тоглы и как ими пользоваться:
1.
Чтобы получить доступ к toggles вам понадобится модифицированное приложение ВКонтакте (напоминаем, что речь идет о Android). Откуда его взять? Есть 2 варианта:
Первый — скачать одну из модификаций приложения с доступом к toggles:
1) VK Equals — мод с небольшим набором доп. возможностей, но частыми обновлениями на новые версии VK и доступом к toggles.
2) VK Tea — мод с целым рядом доп. функций и доступом к toggles. Базируется на версии VK 8.1 и больше не развивается. (Для доступа к toggles необходимо активировать debug в Настройках Tea → Прочее).
Второй — активировать debug-меню самостоятельно с помощью Lucky Patcher.
Данный способ может не сработать без ROOT-прав, но это индивидуально. Так же при выполнении действий без ROOT-прав, приложение VK, в последствии, может работать медленнее.
1) Скачайте приложение Lucky Patcher: скачать;
2) Откройте Lucky Patcher, выдайте ему требуемые разрешения, а так же ROOT-доступ;
3) Откройте боковое меню Lucky Patcher’a и нажмите «Скачать пользовательские патчи«;
4) По окончанию загрузки найдите в списке приложений VK, долгим тапом по нему откройте меню действий и выберите «Пользовательский патч«;
5) В открывшемся списке патчей найдите патч «Hepolise-debug_menu…«, выберите его, нажмите кнопку «Патч«, а затем «Применить» в окне подтверждения.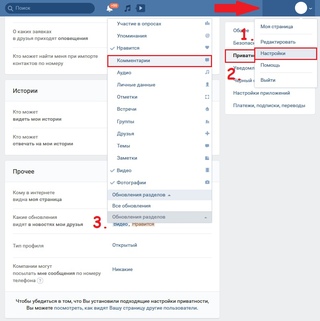
• Скриншоты к инструкции: смотреть.
При обновлении приложения, эти действия придётся повторять!
2.
После того как вы поставили себе модификацию из предложенных выше, перейдите в настройки приложения VK → Debug (Отладка) → toggles. Это и есть скрытые функции ВК!
Не рекомендуем что-либо «трогать» во вкладке General, если вы чётко не знаете, что для чего эти опции.3.
В этом списке что-то выключено (чаще всего это скрытые новинки), а то, что уже добавлено для всех — включено.
Если вы включили или выключили какой-то тогл, для применения результата необходимо перезапустить приложение!
Чтобы сбросить тоглы на первоначальный вариант — выйдете из аккаунта VK, снова войдите в него и перезапустите приложение.—————————————-Канал со списоком полезных и интересных toggles: → ОТКРЫТЬ ←—————————————-
Автор: Telegram-канал VK Leaks | Сливы секретов и новинок ВК
Надеемся, эта информация оказалась полезной!Индукция и эффекторные функции клеток Th27
1. Mosmann TR & Coffman RL
Клетки T H 1 и T H 2: разные модели секреции лимфокинов приводят к различным функциональным свойствам. Анну. Преподобный Иммунол
7, 145–173 (1989). [PubMed] [Google Scholar]
Mosmann TR & Coffman RL
Клетки T H 1 и T H 2: разные модели секреции лимфокинов приводят к различным функциональным свойствам. Анну. Преподобный Иммунол
7, 145–173 (1989). [PubMed] [Google Scholar]
2. Форт ММ и другие. IL-25 индуцирует IL-4, IL-5 и IL-13 и Th3-ассоциированные патологии in vivo . Иммунитет 15, 985–995 (2001). [PubMed] [Google Scholar]
3. Лангриш CL и другие. IL-23 управляет патогенной популяцией Т-клеток, которая вызывает аутоиммунное воспаление. Дж. Эксп. Мед 201, 233–240 (2005). [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Коллс Дж. К. и Линден А. Члены семьи интерлейкина-17 и воспаление. Иммунитет 21, 467–476 (2004). [PubMed] [Google Scholar]
5. Гунималадеви И., Саван Р. и Сакаи М. Идентификация, клонирование и характеристика интерлейкина-17 и его семейства у рыбок данио. Иммунол рыбных моллюсков. 21, 393–403 (2006). [PubMed] [Google Scholar]
6. Liang SC
и другие.
Интерлейкин (IL)-22 и IL-17 коэкспрессируются клетками Th27 и совместно усиливают экспрессию противомикробных пептидов. Дж. Эксп. Мед
203, 2271–2279(2006). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Эксп. Мед
203, 2271–2279(2006). [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Chang SH & Dong C Новый гетеродимерный цитокин, состоящий из IL-17 и IL-17F, регулирует воспалительные реакции. Сотовый рез. 17, 435–440 (2007). [PubMed] [Google Scholar]
8. Liang SC и другие. Гетеродимерный белок IL-17F/A продуцируется клетками Th27 мыши и индуцирует рекрутирование нейтрофилов дыхательных путей. Дж. Иммунол 179, 7791–7799 (2007). [PubMed] [Google Scholar]
9. Чжэн И и другие. Интерлейкин-22, Т Н 17 цитокин, опосредует индуцированное IL-23 кожное воспаление и акантоз. Природа 445, 648–651 (2007). [PubMed] [Google Scholar]
10. Корн Т. и другие. IL-21 инициирует альтернативный путь индукции провоспалительных клеток T H 17. Природа 448, 484–487 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Нуриева Р.
и другие.
Существенная аутокринная регуляция IL-21 в образовании воспалительных Т-клеток. Природа
448, 480–483 (2007). [PubMed] [Академия Google]
[PubMed] [Академия Google]
12. Чжоу Л. и другие. IL-6 программирует дифференцировку клеток T H -17, способствуя последовательному включению путей IL-21 и IL-23. Натура Иммунол. 8, 967–974 (2007). [PubMed] [Google Scholar]
13. Чтанова Т. и другие. Т-фолликулярные хелперные клетки имеют характерный профиль транскрипции, отражающий их роль эффекторных клеток, отличных от Th2/Th3, которые оказывают помощь В-клеткам. Дж. Иммунол 173, 68–78 (2004). [PubMed] [Google Scholar]
14. Ye P и другие. Необходимость передачи сигналов рецептора интерлейкина 17 для экспрессии хемокинов CXC легких и гранулоцитарного колониестимулирующего фактора, рекрутирования нейтрофилов и защиты хозяина. Дж. Эксп. Мед 194, 519–527 (2001). [Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Mangan PR и другие. Трансформирующий фактор роста-β индуцирует развитие линии T H 17. Природа 441, 231–234 (2006). [PubMed] [Google Scholar]
16. Чанг Д.Р.
и другие. CD4 1 Т-клетки опосредуют образование абсцесса при интраабдоминальном сепсисе посредством IL-17-зависимого механизма. Дж. Иммунол
170, 1958–1963 (2003). [PubMed] [Google Scholar]
CD4 1 Т-клетки опосредуют образование абсцесса при интраабдоминальном сепсисе посредством IL-17-зависимого механизма. Дж. Иммунол
170, 1958–1963 (2003). [PubMed] [Google Scholar]
17. Инфанте-Дуарте С., Хортон Х.Ф., Бирн М.С. и Камрадт Т. Микробные липопептиды индуцируют продукцию IL-17 в клетках Th. Дж. Иммунол 165, 6107–6115 (2000). [PubMed] [Академия Google]
18. Хадер С.А. и другие. IL-23 и IL-17 в создании защитного ответа легких CD4 1 Т-клеток после вакцинации и во время заражения Mycobacterium tuberculosis. Натура Иммунол. 8, 369–377 (2007). [PubMed] [Google Scholar]
19. Хуанг В., На Л., Фидель П.Л. и Шварценбергер П. Потребность в интерлейкине-17А для системной защиты хозяина от Candida albicans у мышей. Дж. Заразить. Дис 190, 624–631 (2004). [PubMed] [Google Scholar]
20. ван Билен А.Дж.
и другие.
Стимуляция внутриклеточного бактериального сенсора NOD2 программирует дендритные клетки на стимулирование выработки интерлейкина-17 в Т-клетках памяти человека.
21. Чарльтон Б. и Лафферти К.Дж. Баланс Th2/Th3 при аутоиммунитете. Курс. мнение Иммунол 7, 793–798 (1995). [PubMed] [Google Scholar]
22. Фербер И.А. и другие. Мыши с нарушенным геном IFN-γ подвержены индукции экспериментального аутоиммунного энцефаломиелита (ЭАЭ). Дж. Иммунол 156, 5–7 (1996). [PubMed] [Google Scholar]
23. Бехер Б., Дурелл Б.Г. и Ноэль Р.Дж. Экспериментальный аутоиммунный энцефалит и воспаление в отсутствие интерлейкина-12. Дж. Клин. Вкладывать деньги 110, 493–497 (2002). [Бесплатная статья PMC] [PubMed] [Google Scholar]
24. Cua DJ и другие. Интерлейкин-23, а не интерлейкин-12, является критическим цитокином для аутоиммунного воспаления головного мозга. Природа 421, 744–748 (2003). [PubMed] [Google Scholar]
25. Сато К.
и другие.
Th27 функционирует как подгруппа остеокластогенных хелперных Т-клеток, которая связывает активацию Т-клеток и разрушение кости. Дж. Эксп. Мед
203, 2673–2682 (2006).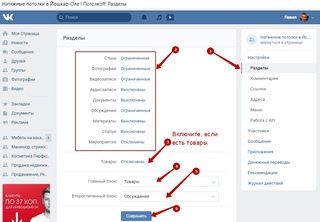
26. Накаэ С., Намбу А., Судо К. и Ивакура Й. Подавление иммунной индукции коллаген-индуцированного артрита у мышей с дефицитом IL-17. Дж. Иммунол 171, 6173–6177 (2003). [PubMed] [Академия Google]
27. Комияма Ю. и другие. IL-17 играет важную роль в развитии экспериментального аутоиммунного энцефаломиелита. Дж. Иммунол 177, 566–573 (2006). [PubMed] [Google Scholar]
28. Шабо М. и другие. Человеческий интерлейкин-17: провоспалительный цитокин Т-клеток, продуцируемый ревматоидной синовиальной оболочкой. Ревмирующий артрит. 42, 963–970 (1999). [PubMed] [Google Scholar]
29. Замок C и другие. Анализ генных микрочипов очагов рассеянного склероза дает новые мишени, подтвержденные при аутоиммунном энцефаломиелите. Природа Мед. 8, 500–508 (2002). [PubMed] [Академия Google]
30. Фуджино С. и другие. Повышенная экспрессия интерлейкина 17 при воспалительных заболеваниях кишечника. кишки 52, 65–70 (2003). [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Wilson NJ
и другие.
Развитие, цитокиновый профиль и функция человеческих Т-хелперных клеток, продуцирующих интерлейкин 17. Натура Иммунол. 8, 950–957 (2007). [PubMed] [Google Scholar]
Wilson NJ
и другие.
Развитие, цитокиновый профиль и функция человеческих Т-хелперных клеток, продуцирующих интерлейкин 17. Натура Иммунол. 8, 950–957 (2007). [PubMed] [Google Scholar]
32. Кебир Х. и другие. Лимфоциты T H 17 человека способствуют нарушению гематоэнцефалического барьера и воспалению центральной нервной системы. Природа Мед. 13, 1173–1175 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Бен-Нун А., Векерле Х. и Коэн И.Р. Быстрое выделение клонируемых антиген-специфических линий Т-лимфоцитов, способных вызывать аутоиммунный энцефаломиелит. Евро. Дж. Иммунол 11, 195–199 (1981). [PubMed] [Google Scholar]
34. Lohr J, Knoechel B, Wang JJ, Villarino AV & Abbas AK Роль IL-17 и регуляторных Т-лимфоцитов в системном аутоиммунном заболевании. Дж. Эксп. Мед 203, 2785–2791 (2006). [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Zenewicz LA
и другие.
Интерлейкин-22, но не интерлейкин-17, обеспечивает защиту гепатоцитов при остром воспалении печени. Иммунитет
27, 647–659(2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Иммунитет
27, 647–659(2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM & Stockinger B TGFβ в контексте воспалительной цитокиновой среды поддерживает дифференцировку de novo Т-клеток, продуцирующих IL-17. Иммунитет 24, 179–189 (2006). [PubMed] [Google Scholar]
37. Беттелли Э. и другие. Реципрокные пути развития образования патогенных эффекторных Th27 и регуляторных Т-клеток. Природа 441, 235–238 (2006). [PubMed] [Академия Google]
38. Кулкарни А.Б. и другие. Нулевая мутация трансформирующего фактора роста бета 1 у мышей вызывает чрезмерную воспалительную реакцию и раннюю смерть. проц. Натл акад. науч. США 90, 770–774 (1993). [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Veldhoen M, Hocking RJ, Flavell RA & Stockinger B
Сигналы, опосредованные трансформирующим фактором роста-бета, инициируют аутоиммунный энцефаломиелит, но для поддержания болезни необходимо хроническое воспаление. Натура Иммунол. 7, 1151–1156 (2006). [PubMed] [Академия Google]
7, 1151–1156 (2006). [PubMed] [Академия Google]
40. Ли М.О., Ван Ю.Ю. и Флавелл Р.Т. Производимый клетками трансформирующий фактор роста-β1 контролирует толерантность Т-клеток и регулирует дифференцировку Th2- и Th27-клеток. Иммунитет 26, 579–591 (2007). [PubMed] [Google Scholar]
41. Акоста-Родригес EV и другие. Поверхностный фенотип и антигенная специфичность человеческих Т-хелперных клеток памяти, продуцирующих интерлейкин 17. Натура Иммунол. 8, 639–646 (2007). [PubMed] [Google Scholar]
42. Сато В., Аранами Т. и Ямамура Т. Новшество: клетки Th27 человека идентифицированы как несущие CCR2 1 CCR5 2 фенотип. Дж. Иммунол 178, 7525–7529 (2007). [PubMed] [Google Scholar]
43. Acosta-Rodriguez EV, Napolitani G, Lanzavecchia A & Sallusto F Интерлейкины 1β и 6, но не трансформирующий фактор роста-β, необходимы для дифференцировки Т-хелперных клеток человека, продуцирующих интерлейкин 17. Природа Иммунол 8, 942–949 (2007). [PubMed] [Google Scholar]
44. Evans HG, Suddason T, Jackson I, Taams LS & Lord GM
Оптимальная индукция Т-хелперов 17 у людей требует лигирования Т-клеточного рецептора в контексте моноцитов, активируемых Толл-подобным рецептором. проц. Натл акад. науч. США
104, 17034–17039(2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Evans HG, Suddason T, Jackson I, Taams LS & Lord GM
Оптимальная индукция Т-хелперов 17 у людей требует лигирования Т-клеточного рецептора в контексте моноцитов, активируемых Толл-подобным рецептором. проц. Натл акад. науч. США
104, 17034–17039(2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Chen Z, Tato CM, Muul L, Laurence A & O’Shea JJ Четкая регуляция интерлейкина-17 в Т-хелперных лимфоцитах человека. Ревмирующий артрит. 56, 2936–2946 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Мерфи К.М. и Райнер С.Л. Линейные решения Т-хелперов. Природа Преподобный Иммунол 2, 933–944 (2002). [PubMed] [Google Scholar]
47. Sutton C, Brereton C, Keogh B, Mills KH & Lavelle EC Решающая роль интерлейкина (IL)-1 в индукции IL-17-продуцирующих Т-клеток, которые опосредуют аутоиммунный энцефаломиелит. Дж. Эксп. Мед 203, 1685–169 гг.1 (2006). [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Штумхофер Дж. С.
и другие.
Интерлейкины 27 и 6 индуцируют STAT3-опосредованную продукцию интерлейкина 10 Т-клетками. Nature Immunol. 8, 1363–1371 (2007). [PubMed] [Google Scholar]
Nature Immunol. 8, 1363–1371 (2007). [PubMed] [Google Scholar]
49. McGeachy MJ и другие. TGF-β и IL-6 стимулируют продукцию IL-17 и IL-10 Т-клетками и сдерживают клеточно-опосредованную патологию T H -17. Натура Иммунол. 8, 1390–1397 (2007). [PubMed] [Google Scholar]
50. Оппманн Б. и другие. Роман р19белок взаимодействует с IL-12p40 с образованием цитокина IL-23 с биологической активностью, сходной с IL-12 и отличной от нее. Иммунитет 13, 715–725 (2000). [PubMed] [Google Scholar]
51. Kastelein RA, Hunter CA & Cua DJ Открытие и биология IL-23 и IL-27: родственные, но функционально разные регуляторы воспаления. Анну. Преподобный Иммунол 25, 221–242 (2007). [PubMed] [Google Scholar]
52. Парэм С. и другие. Рецептор гетеродимерного цитокина IL-23 состоит из IL-12Rb1 и новой субъединицы рецептора цитокинов, IL-23R. Дж. Иммунол 168, 5699–5708 (2002). [PubMed] [Google Scholar]
53. Uhlig HH
и другие.
Дифференциальная активность ИЛ-12 и ИЛ-23 при патологии слизистых оболочек и системной врожденной иммунной патологии. Иммунитет
25, 309–318 (2006). [PubMed] [Google Scholar]
Иммунитет
25, 309–318 (2006). [PubMed] [Google Scholar]
54. Дюрр Р.Х. и другие. Полногеномное исследование ассоциации идентифицирует IL23R как ген воспалительного заболевания кишечника. Наука 314, 1461–1463 (2006). [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Park H и другие. Отдельная линия Т-клеток CD4 регулирует воспаление тканей, продуцируя интерлейкин 17. Nature Immunol. 6, 1133–1141 (2005). [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Харрингтон LE и другие. Эффекторные Т-клетки CD4 + , продуцирующие интерлейкин 17, развиваются в линии, отличной от линий Т-хелперов 1 и 2 типов. Натура Иммунол. 6, 1123–1132 (2005). [PubMed] [Google Scholar]
57. Чен З. и другие. Селективная регуляторная функция Socs3 в формировании секретирующих IL-17 Т-клеток. проц. Натл акад. науч. США 103, 8137–8142 (2006). [Бесплатная статья PMC] [PubMed] [Google Scholar]
58. Yang XO
и другие.
STAT3 регулирует опосредованную цитокинами генерацию воспалительных хелперных Т-клеток.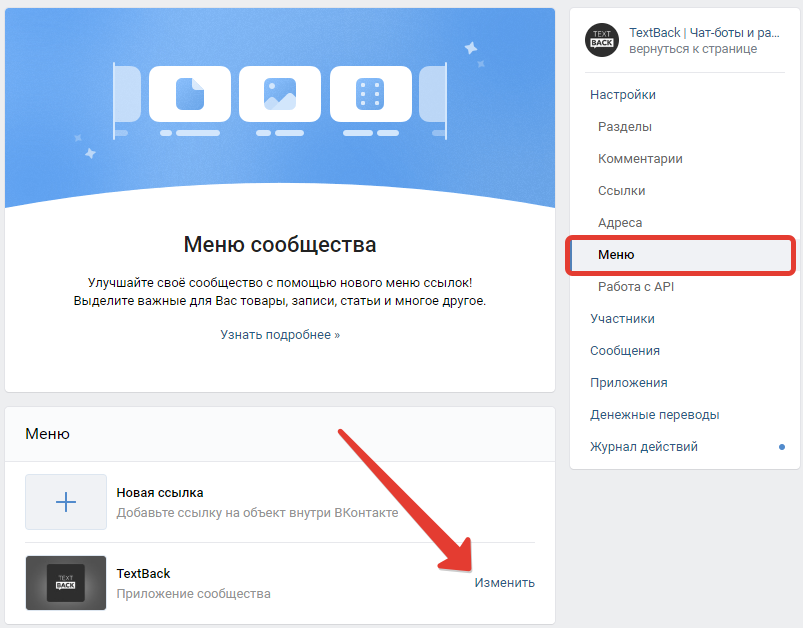 Дж. Биол. Химия
282, 9358–9363 (2007). [PubMed] [Google Scholar]
Дж. Биол. Химия
282, 9358–9363 (2007). [PubMed] [Google Scholar]
59. Иванов И.И. и другие. Орфанный ядерный рецептор RORct направляет программу дифференцировки провоспалительных IL-17 1 Т-хелперных клеток. Клетка 126, 1121–1133 (2006). [PubMed] [Google Scholar]
60. Ян XO и другие. Дифференцировка линии Т-хелперов 17 программируется орфанными ядерными рецепторами RORa и RORc. Иммунитет 28, 29–39 (2008). [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Лоуренс А. и другие. Передача сигналов интерлейкина-2 через STAT5 ограничивает образование Т-хелперов 17. Иммунитет 26, 371–381 (2007). [PubMed] [Академия Google]
62. Муцида Д. и другие. Реципрокная T H 17 и регуляторная дифференцировка Т-клеток, опосредованная ретиноевой кислотой. Наука 317, 256–260 (2007). [PubMed] [Google Scholar]
63. Антов А., Ян Л., Виг М., Балтимор Д. и Ван Парийс Л.
Существенная роль для передачи сигналов STAT5 в CD25 1 CD4 1 Регуляторный гомеостаз Т-клеток и поддержание толерантности к себе. Дж. Иммунол
171, 3435–3441 (2003). [PubMed] [Google Scholar]
Дж. Иммунол
171, 3435–3441 (2003). [PubMed] [Google Scholar]
64. Кумбс Дж.Л. и другие. Функционально специализированная популяция слизистой CD103 1 ДК индуцируют регуляторные Т-клетки Foxp3 1 через механизм, зависящий от TGF-β и ретиноевой кислоты. Дж. Эксп. Мед 204, 1757–1764 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Чжоу Л. и другие. TGF-β-индуцированный Foxp3 ингибирует дифференцировку клеток T H 17, противодействуя функции RORct. Природа 453, 236–240 (2008). [Бесплатная статья PMC] [PubMed] [Google Scholar]
66. Du J, Huang C, Zhou B & Ziegler SF Изоформ-специфическое ингибирование RORα-опосредованной активации транскрипции человеческим FOXP3. Дж. Иммунол 180, 4785–4792 (2008). [PubMed] [Google Scholar]
67. Гэвин М.А. и другие. Foxp3-зависимая программа регуляторной дифференцировки Т-клеток. Природа 445, 771–775 (2007). [PubMed] [Google Scholar]
68. Уильямс Л.М. и Руденски А.Ю.
Поддержание Foxp3-зависимой программы развития в зрелых регуляторных Т-клетках требует постоянной экспрессии Foxp3. Натура Иммунол. 8, 277–284 (2007). [PubMed] [Google Scholar]
Натура Иммунол. 8, 277–284 (2007). [PubMed] [Google Scholar]
69. Кляйншек М.А. и другие. IL-25 регулирует функцию Th27 при аутоиммунном воспалении. Дж. Эксп. Мед 204, 161–170 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
70. Доска М и другие. Интерлейкин 27 ограничивает аутоиммунный энцефаломиелит, подавляя развитие Т-клеток, продуцирующих интерлейкин 17. Натура Иммунол. 7, 929–936 (2006). [PubMed] [Google Scholar]
71. Штумхофер Дж. С. и другие. Интерлейкин 27 отрицательно регулирует развитие интерлейкина 17-продуцирующих Т-хелперов при хроническом воспалении центральной нервной системы. Натура Иммунол. 7, 937–945 (2006). [PubMed] [Google Scholar]
72. Авасти А. и другие. Доминирующая функция интерлейкина 27 в создании противовоспалительных Т-клеток, продуцирующих интерлейкин 10. Натура Иммунол. 8, 1380–1389 гг.(2007). [PubMed] [Google Scholar]
73. Лю С.Дж.
и другие.
Индукция отдельного подмножества CD8 Tnc17 путем трансформации фактора роста-β и интерлейкина-6. Дж. Лейкок. Биол
82, 354–360 (2007). [PubMed] [Google Scholar]
Дж. Лейкок. Биол
82, 354–360 (2007). [PubMed] [Google Scholar]
74. Локхарт Э., Грин А.М. и Флинн Д.Л. В продукции IL-17 доминируют Т-клетки cd, а не Т-клетки CD4 во время инфекции Mycobacterium tuberculosis. Дж. Иммунол 177, 4662–4669 (2006). [PubMed] [Google Scholar]
75. Ferretti S, Bonneau O, Dubois GR, Jones CE & Trifilieff A ИЛ-17, продуцируемый лимфоцитами и нейтрофилами, необходим для липополисахарид-индуцированной нейтрофилии дыхательных путей: ИЛ-15 как возможный триггер. Дж. Иммунол 170, 2106–2112 (2003). [PubMed] [Академия Google]
76. Молет С и другие. Уровень IL-17 увеличивается в дыхательных путях, страдающих астмой, и вызывает образование цитокинов фибробластами бронхов человека. Дж. Аллергия Клин. Иммунол 108, 430–438 (2001). [PubMed] [Google Scholar]
77. Чжоу К., Деста Т., Фентон М., Грейвс Д.Т. и Амар С.
Профилирование цитокинов макрофагов, подвергшихся воздействию Porphyromonas gingivalis, его липополисахарида или его белка FimA. Заразить. Иммунная
73, 935–943 (2005). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Иммунная
73, 935–943 (2005). [Бесплатная статья PMC] [PubMed] [Google Scholar]
78. Yao Z и другие. Вирус герпеса Saimiri кодирует новый цитокин IL-17, который связывается с новым цитокиновым рецептором. Иммунитет 3, 811–821 (1995). [PubMed] [Google Scholar]
79. Kuestner RE и другие. Идентификация молекулы IL-17RC, родственной рецептору IL-17, в качестве рецептора для IL-17F. Дж. Иммунол 179, 5462–5473 (2007). [Бесплатная статья PMC] [PubMed] [Google Scholar]
80. Toy D и другие. Передний край: сигналы интерлейкина 17 через гетеромерный рецепторный комплекс. Дж. Иммунол 177, 36–39 (2006). [PubMed] [Google Scholar]
81. Фоссиес Ф. и другие. Т-клеточный интерлейкин-17 побуждает стромальные клетки продуцировать провоспалительные и гемопоэтические цитокины. Дж. Эксп. Мед 183, 2593–2603 (1996). [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Мартель-Пеллетье Дж., Мино Ф., Йованович Д., Ди Баттиста Дж.А. и Пеллетье Дж.П.
Митоген-активируемая протеинкиназа и ядерный фактор kB вместе регулируют индуцированную интерлейкином-17 продукцию оксида азота в хондроцитах человека при остеоартрите: возможная роль трансактивирующего фактора митоген-активируемая протеинкиназа-активируемая протеинкиназа (MAPKAPK).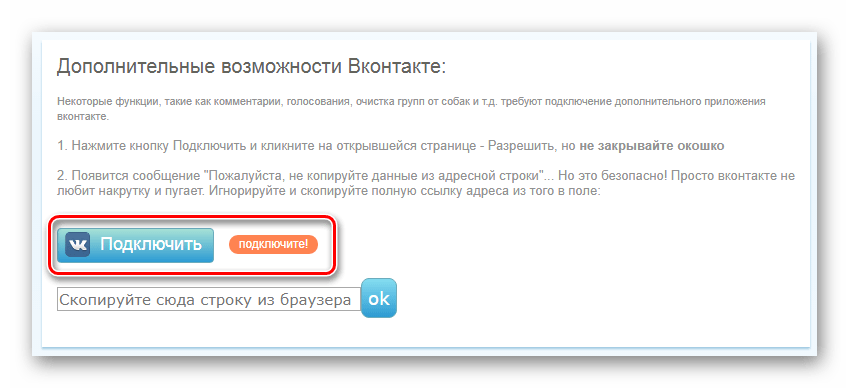 Ревмирующий артрит. 42, 2399–2409 (1999). [PubMed] [Google Scholar]
Ревмирующий артрит. 42, 2399–2409 (1999). [PubMed] [Google Scholar]
83. Старк М.А. и другие. Фагоцитоз апоптотических нейтрофилов регулирует гранулопоэз через IL-23 и IL-17. Иммунитет 22, 285–294 (2005). [PubMed] [Google Scholar]
84. Старнес Т. и другие. Передовой опыт: IL-17F, новый цитокин, селективно экспрессируемый в активированных Т-клетках и моноцитах, регулирует ангиогенез и продукцию цитокинов эндотелиальными клетками. Дж. Иммунол 167, 4137–4140 (2001). [PubMed] [Google Scholar]
85. Hymowitz SG и другие. IL-17 принимают складку цистинового узла: структура и активность нового цитокина, IL-17F, и значение для связывания с рецептором. EMBO J. 20, 5332–5341 (2001). [Бесплатная статья PMC] [PubMed] [Google Scholar]
86. Херст С.Д. и другие. Новые члены семейства IL-17 способствуют ответам Th2 или Th3 в легких: in vivo функция нового цитокина IL-25. Дж. Иммунол 169, 443–453 (2002). [PubMed] [Google Scholar]
87. Wolk K & Sabat R
Интерлейкин-22: новый цитокин, полученный из Т- и NK-клеток, который регулирует биологию тканевых клеток. Cytokine Growth Factor Rev. 17, 367–380 (2006). [PubMed] [Google Scholar]
Cytokine Growth Factor Rev. 17, 367–380 (2006). [PubMed] [Google Scholar]
88. Котенко С.В. и другие. Идентификация функционального рецепторного комплекса интерлейкина-22 (IL-22): цепь IL-10R2 (IL-10Rb) является общей цепью как IL-10, так и IL-22 (индуцируемый Т-клетками родственный IL-10 фактор, IL-TIF) рецепторные комплексы. Дж. Биол. Химия 276, 2725–2732 (2001). [PubMed] [Академия Google]
89. Мур К.В., де Ваал Малефит Р., Коффман Р.Л. и О’Гарра А. Интерлейкин-10 и рецептор интерлейкина-10. Анну. Преподобный Иммунол 19, 683–765 (2001). [PubMed] [Google Scholar]
90. Волк К. и другие. ИЛ-22 повышает врожденный иммунитет тканей. Иммунитет 21, 241–254 (2004). [PubMed] [Google Scholar]
91. Дюмутье Л., Ван Руст Э., Коло Д. и Рено Дж. К. Связанный с интерлейкином-10 человека индуцируемый фактор Т-клеток: молекулярное клонирование и функциональная характеристика как фактора, стимулирующего гепатоциты. проц. Натл акад. науч. США 97, 10144–10149 (2000). [Бесплатная статья PMC] [PubMed] [Google Scholar]
92. Пэрриш-Новак Дж.
и другие.
Интерлейкин 21 и его рецептор участвуют в размножении NK-клеток и регуляции функции лимфоцитов. Природа
408, 57–63 (2000). [PubMed] [Google Scholar]
Пэрриш-Новак Дж.
и другие.
Интерлейкин 21 и его рецептор участвуют в размножении NK-клеток и регуляции функции лимфоцитов. Природа
408, 57–63 (2000). [PubMed] [Google Scholar]
93. Леонард В.Дж. и Спольски Р. Интерлейкин-21: модулятор лимфоидной пролиферации, апоптоза и дифференцировки. Природа Преподобный Иммунол 5, 688–698 (2005). [PubMed] [Google Scholar]
94. Такешита Т. и другие. Клонирование гамма-цепи рецептора IL-2 человека. Наука 257, 379–382 (1992). [PubMed] [Google Scholar]
95. Цзэн Р. и другие. Синергизм IL-21 и IL-15 в регуляции размножения и функции CD8 + Т-клеток. Дж. Эксп. Мед 201, 139–148 (2005). [Бесплатная статья PMC] [PubMed] [Google Scholar]
96. Spolski R & Leonard WJ Интерлейкин-21: основная биология и значение для рака и аутоиммунитета. Анну. Преподобный Иммунол 26, 57–79 (2008). [PubMed] [Google Scholar]
97. Coquet JM
и другие.
IL-21 продуцируется NKT-клетками и модулирует активацию NKT-клеток и продукцию цитокинов. Дж. Иммунол
178, 2827–2834 (2007). [PubMed] [Академия Google]
Дж. Иммунол
178, 2827–2834 (2007). [PubMed] [Академия Google]
98. Пеллетье М., Бушар А. и Жирар Д. In vivo и in vitro роль IL-21 в воспалении. Дж. Иммунол 173, 7521–7530 (2004). [PubMed] [Google Scholar]
99. Манель Н., Унутмаз Д. и Литтман Д.Р. Дифференцировка клеток T(H)-17 человека требует трансформирующего фактора роста-бета и индукции ядерного рецептора RORgamma. Нац. Иммунол предварительная онлайн-публикация doi:10.1038/ni.1610 (4 Может 2008). [Бесплатная статья PMC] [PubMed] [Google Scholar]
100. Ян Л. и другие. IL-21 и TGF-β необходимы для дифференцировки T 9 человека.0003 H 17 ячеек. Природа предварительная онлайн-публикация doi:10.1038/nature07021 (11 Может 2008). [PMC free article] [PubMed] [Google Scholar]
Полуавтоматическая этикетировочная машина для круглых бутылок VK-T801
Перейти к содержимомуВведение
Полуавтоматическая этикетировочная машина предназначена для маркировки круглых бутылок диаметром от 25 до 25 мм. до 160мм. Он оснащен цилиндром для разглаживания этикетки с максимальной производительностью 40 штук в минуту.
до 160мм. Он оснащен цилиндром для разглаживания этикетки с максимальной производительностью 40 штук в минуту.
Эта машина представляет собой полуавтоматическую этикетировочную машину, предназначенную для этикетирования цилиндрических и круглых бутылок с малым конусом. Он может выполнять маркировку на всю неделю или полнедели, круглую обратную этикетку и регулируемую обратную этикетку. Он позволяет наносить одинарную маркировку, маркировку с фиксированной точкой или двойную маркировку.
Этикетировочная машина ВК-Т801 может выполнять этикетирование одним кругом и полукругом, а также двойное этикетирование с обеих сторон изделия. Расстояние между передней и задней этикетками регулируется, и метод регулировки также прост. Он находит широкое применение в пищевой, косметической, химической, винодельческой, фармацевтической и других отраслях промышленности.
1. Он может выполнять обтекание одной этикеткой вокруг этикетки, две этикетки на лицевой и оборотной стороне, а также ориентацию маркировки.
2. Можно добавить принтер даты для печати даты производства, партии партии и даты истечения срока годности при маркировке.
3. Доступна поддержка настройки.
Просмотр видеоОписаниеДетали машиныРабочее видеоСвяжитесь с намиСопутствующие товары
VK-T801 имеет дополнительные функции для добавления опций:
1. Добавьте функцию позиционной маркировки, чтобы этикетку можно было прикрепить к фиксированному положению на вашем продукте.
2. Оснащенный кодировщиком или струйным принтером, номер производственной партии, дата производства, дата вступления в силу и другая информация четко печатаются во время маркировки. Это позволяет одновременно выполнять кодирование и маркировку, повышая эффективность.
Способ регулировки VK-T801 прост и требует только регулировки высоты прижимного ролика и ширины отверстия, в которое помещается изделие. Процесс настройки занимает менее 5 минут, а точность маркировки высокая. Ошибки обнаружить невооруженным глазом сложно.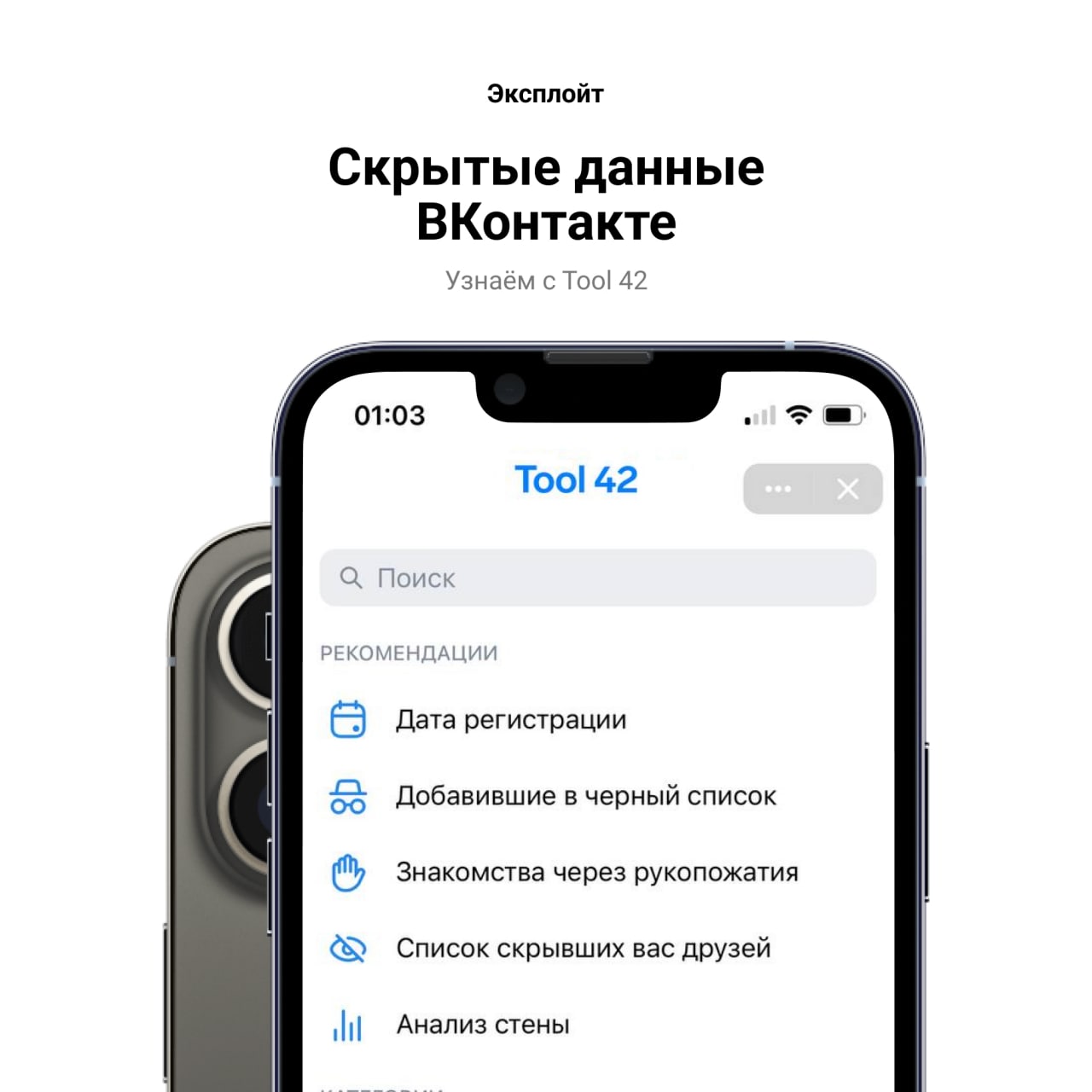 VK-T801 занимает площадь около 0,22 куб. Он также поддерживает пользовательские этикетировочные машины в зависимости от требований к продукту.
VK-T801 занимает площадь около 0,22 куб. Он также поддерживает пользовательские этикетировочные машины в зависимости от требований к продукту.
| Параметр | Дата |
| Этикетка со спецификацией | Клейкая наклейка, прозрачная или непрозрачная |
| Допуск маркировки | ±0,5 мм |
| 15~30 | |
| Размер бутылки костюма ( мм) | Ø15~Ø150;можно настроить |
| Размер этикетки (мм) | Д:20~290;Ш(В):15~130 |
| Размер машины(Д*Ш*В) | ≈960*560*540(мм) |
| Размер упаковки (Д*Ш*В) | ≈1020*660*740 (мм) |
| Напряжение | 220 В/50 (60) Гц; можно настроить | Мощность | 120 Вт |
| Вес (кг) | ≈45,0 |
| Вес (кг) | ≈67,5 |
| Рулон этикеток | ID: Ø76 мм; Внешний диаметр:≤260 мм |
| Подача воздуха | 0,4~0,6 МПа |
Требования к производству этикеток
1. Зазор между этикеткой и этикеткой должен быть 2-3мм.
Зазор между этикеткой и этикеткой должен быть 2-3мм.
2. Расстояние между этикеткой и краем нижней бумаги должно быть 2 мм.
3. Нижняя бумага этикетки изготовлена из пергамина, который обладает хорошей прочностью и не дает ей сломаться (чтобы не порезать нижнюю бумагу).
4. Внутренний диаметр сердечника должен быть 76 мм, а внешний диаметр должен быть менее 300 мм. Этикетки должны располагаться в один ряд.
Приведенные выше требования к производству этикеток должны быть включены в ваш продукт. Подробности смотрите в результатах общения с нашими инженерами.
Применение
Примеры применения: бутылка соуса чили, круглая бутылка жидкости для перорального применения, бутылка Celine, ксилит, маркировка бутылок с шампунем, двойная этикетка для вина, маркировка с фиксированной точкой для вина, позитивная маркировка и т. д.
Рабочий процесс
Нажмите переключатель, поместив изделие в указанное положение.
Машина зажмет продукт и вытянет этикетку. Колесо в верхней части машины будет прижимать этикетку к продукту, а затем вращать ее до тех пор, пока маркировка не будет завершена.
Колесо в верхней части машины будет прижимать этикетку к продукту, а затем вращать ее до тех пор, пока маркировка не будет завершена.
Отпустите продукт, и машина автоматически восстановится.
Процесс маркировки завершен.
Характеристики
- Мощные функции, включая маркировку одной и двумя этикетками, можно произвольно переключать. Расстояние между передней и задней двойными этикетками можно гибко регулировать. Машина может быть оснащена функцией позиционирования по окружности, позволяющей установить положение маркировки на поверхности по окружности.
- Процесс эксплуатации прост и удобен. Автоматическое прижатие цилиндра входит в стандартную комплектацию, что снижает нагрузку на оператора при загрузке и разгрузке и повышает эффективность этикетирования.
- Дополнительный ленточный принтер и струйный принтер для согласования цветов доступны для одновременной маркировки и печати номеров производственных партий и другой информации, что сокращает процесс упаковки и повышает эффективность производства.

- Предусмотрена опора для маркировки конических бутылок с ручкой регулировки конусности для легкой регулировки в соответствии с требованиями к маркировке конических бутылок.
- Превосходное качество этикетирования благодаря двойному позиционированию для позиционирования продукта и коррекции отклонения этикетки, что обеспечивает высокую степень перекрытия между головной и задней частью этикетки. Нет морщин или пузырей, что повышает эффективность и обеспечивает качество.
- Возможна простая регулировка с переключением различных функций одной кнопкой, что позволяет быстро переключать маркировку для разных этикеток продукта, что повышает эффективность.
- Оборудование безопасно и гигиенично, соответствует производственным требованиям GMP. Основными используемыми материалами являются нержавеющая сталь и высококачественный алюминиевый сплав, что делает его прочным и долговечным.
- Стабильная производительность достигается за счет использования ПЛК + сенсорный экран + электронная система управления с оптоволоконным датчиком, поддерживающая долгие часы работы.