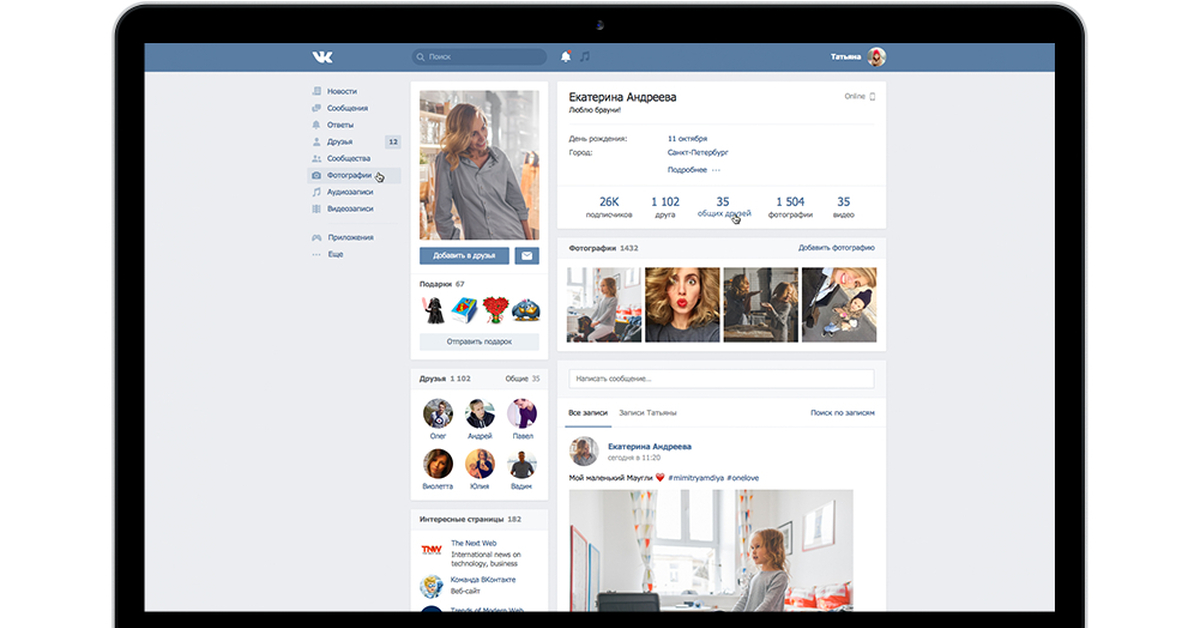
QR код для обновления ВК в 2021 году
Популярная социальная сеть Вконтакте выпустила специальное обновление которое в корне меняет внешний вид приложения. Оно стало в разы удобнее и быстрее, а главное появились функции которые пользователи так долго ждали. Для перехода на новый дизайн, требуется просканировать специальный QR код который автоматически запустит ссылку обновления ВК.
Как получить QR код обновления ВК в 2021 году?
Получить специальный QR код достаточно просто если следовать простой инструкции. Обычным пользователям на данный момент не расшаривают доступ, а только избранным, кто готов участвовать в бета-тестировании дизайна. Если вам не терпится опробовать новый внешний вид, то для активации делаем следующее:
Инструкция без QR кода
- Установите последнюю версию приложения Вконтакте
- Приложение начнет обновление и пользователь должен увидеть следующее:
- После успешного выполненеия процедуры, мы увидим такое сообщение:
- Нажимаем кнопку Continue (Продолжить) и радуемся новому дизайну.

Инструкция по обновлению через QR код
Данный метод не сильно будет отличаться от предыдущего и имеет под собой общую основу взаимодействия.
- Устанавливаем последнюю версию приложения по ссылкам ниже:
- Открываем приложение и переходим в раздел «Истории».
- Теперь наводим камеру на QR-код который расположен ниже:
- При успешном сканировании Вы сразу увидите следующий процесс:
- Новый дизайн будет установлен в течении одной минуты.
Как вы можете видеть, инструкция достаточно простая и состоит всего из нескольких шагов. Ниже мы хотим рассказать про затруднения у некоторых пользователей которые вызваны после обновления. Но касаются они далеко не всех.
Новый QR код обновления
Следить за всеми новшествами и вовремя получать обновления нужно через специальный раздел на сайте: https://m.vk.com/up
В данном разделе размещаются актуальные обновления и возможные BETA-релизы для установки. Закачка производится путём сканирования QR-кода как и в обычной версии.
Не удалось обновить приложение ВК через QR код — что делать?
В данном случае следует понять на каком шаге наши действия были неверны. Например не все обновляют приложение до последней версии перед установкой нового дизайна. В 2021 году вышло крупный апдейт который исправляет множество проблем при загрузке библиотек с удалённого сервера.
Наши действия при ошибкам установки программы будут следующие:
- Полностью удалите VK и установите заново именно с официальных источников
- Используйте актуальный QR-код 2021 года который мы разместили выше.
- Попробуйте освободить больше системной памяти и очистить кеш приложения.
- Оптимизируйте качество интернет-соединения и заряд аккумулятора вашего дейваса.
Мы надеемся, что у вас получить успешно произвести все действия. Оставьте комментарий если что-то пошло не так и мы поможем решить проблему.
Обновляем приложение вк на андроиде. Вконтакте выпустило обновленное мобильное приложение Новое обновление вконтакте сентябрь
Сегодня 27.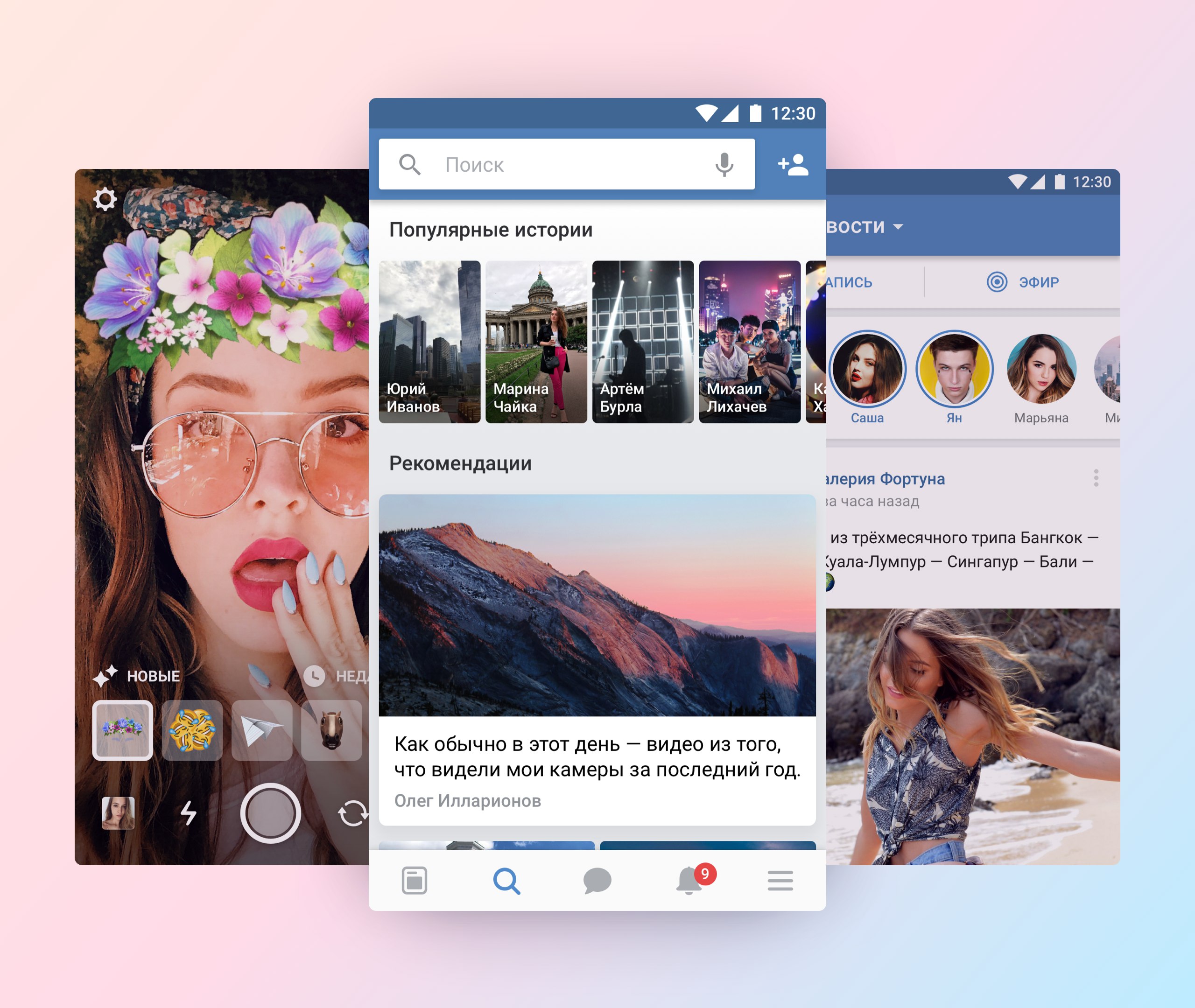 09.2017 я с удивлением обнаружил, что в моём Айфоне, да и на других телефонах с ОС Андроид, появилась возможность обновить . Обновление Мобильного ВК приходиться принимать как данность, резкий переход к новому виду.
09.2017 я с удивлением обнаружил, что в моём Айфоне, да и на других телефонах с ОС Андроид, появилась возможность обновить . Обновление Мобильного ВК приходиться принимать как данность, резкий переход к новому виду.
Быстрая навигация:
Новый вид мобильного приложения VK.
На первый взгляд, очень интересно, что-то новое! Но после нескольких секунд, когда не смог найти в меню вкдадку «Сообщения», меня смутило и я вспомнил все эти дрявые сервисы от маил ру . Конечно же, ВК уже не Павла Дурова. В моей голове прокатилась волна негодования с лозунгами, типа: «Верните Дурова!»…
Что нового и хорошего?
А хорошее действительно есть, как и кто бы чего не говорил. Ведь всё новое всегда воспринимается в штыки.
Новое меню (Удобнее с большим экраном)
Все самые важные функции вынесены на нижнюю панель, остальные пункты меню находятся теперь в правом нижнем углу. С таким интерфейсом действительно стало удобнее и добраться к примеру до сообщений, можно в 1 клик.
Теперь обладателям смартфонов с большим экраном, будет намного удобнее пользоваться приложением. Ранее кнопка меню находилась вверху и для того чтобы ей воспользоваться нужна была вторая рука или неимоверные выкрутасы с одной.
Теперь это самое меню находится внизу и его легко можно кликнуть большим пальцем.
Ответы «Колокольчик»
Теперь всё самое важное, друзья, лайки и репосты, всё собрано в одном пункте. Там же есть настройки уведомлений, теперь можно выполнить тонкую настройку уведомлений, под себя. Оставить только то что, действительно важно для вас.
Улучшенный поиск
В новой версии для поиска используется единое поле ввода, здесь вы сможете найти человека, группу, записи, видео и музыку. Поиск стал более интерактивным, на передний план алгоритм, выводит информацию на основе предпочтений пользователя.
Для отключения безопасного поиска, по прежнему нужно переключаться в браузере (См. ).
Менее значимые изменения нового ВК.
Как только я поставил первый , как заметил что он стал красным, сразу же напомнило Инстаграм. Ранее при нажатии, он приобретал более тёмный, голубой цвет. Что сразу понравилось, так это очень акуратно выполнные элементы под новостями, лайки, сообщения, и просмотры. Теперь смотрятся более гармонично и стильно.
Кстати музыка Вконтакте теперь имеет лимит на прослушивание, как обойти такое нововведение (См. ).
Как вам новшества?
Сейчас пока рано давать однозначный ответ, нужно время и это время может заставить нас свыкнуться с новым дизайном и функционалом. Приветственные картинки, рассказывающие о новых фичах, выполнены в отвратительном стиле, где тот, простой дизайн, привык к нему. Сейчас же только добавить бежевый фон и будет аля Одноклассники…
Наверняка в ближайшее время, будет много споров, лучше или хуже, но нам придётся схавать новую обнову, редизайн 2017.
Что касается изменений, на первый взгляд можно заметить что функция сообщений вынесена из основного меню в отдельную иконку нижнего трея, напомнило WhatsApp. Конечно если брать все изменения, то ВК стал похож больше на Facebook и немного на Instagram.
Конечно если брать все изменения, то ВК стал похож больше на Facebook и немного на Instagram.
На чтение 3 мин.
Очень просто можно обновить вк на телефоне андроид. При этом процедура является обязательной, потому как устаревшие версии со временем могут плохо функционировать.
Обновлять программу на телефоне нужно для того, чтобы пользоваться всеми новыми функциями, ведь разработчики постоянно корректируют сайт, создавая что-то новое и совершенное для более удобного пользования .
Новая версия
Новая версия программы, которую каждый может абсолютно бесплатно скачать на свой телефон, может похвастаться переписанным кодом, кроме того, здесь полностью обновлен дизайн. Отметить можно то, что заголовки раздела стали синего цвета.
И это еще далеко не все «новинки» сайта. Вверху раздела «новости», можно воспользоваться вкладками «Поиск» и «Рекомендации». Вкладкой «комментарии» можно воспользоваться в отделе «Ответы». Ранее же все это можно было делать только при помощи свайпа экрана в правой части.
Профили пользователей также расширились, на них отображается информация вся та же, что и на веб-версии . Изменения коснулись и режима прослушивания музыки, в аудио-плеере отныне есть тексты песен и обложки альбомов.
Обновление
Крайне просто загрузить обновления на андроид. Как обновить вк на телефоне андроид:
- Для этого нужно зайти в стандартное приложение всех андроид смартфонов – Google Play. В этом магазине программ и приложений удобнее всего скачивать обновления.
- Сверху слева нужно нажать на меню и выбрать вкладку «Мои приложения».
- Далее будет видно, какие приложения установлены на телефоне, а в самом верху будут обозначены те, которые требуют обновления.
- Ищем среди них приложение Вк и нажимаем «обновить».
- Приложение выведет на экран уведомление о том, что оно будет иметь доступ к идентификационным данным, нужно нажать кнопку «принять».
- После этого обновление начнется скачиваться.
- Программа на этом этапе обновляется автоматически.
Если используется обычный интернет-трафик, то приложение может уведомить о том, что размер скачиваемого файла слишком большой. Это нужно для того чтобы пользователь знал, что возможна задержка или переплата за трафик. Лучше всего обновлять Вк при подключенном Wi-Fi, это позволит сэкономить денежные средства на трафике.
Возможно, что на телефоне осталось мало места. В этом случае приложение также выведет на экран уведомление о том, что загрузить обновление не удалось.
Тогда необходимо будет очистить место на телефоне, для того чтобы обновление все-таки установилось. Удалить можно игры, музыку или фото. Удаляются лишние файлы с внутренней памяти телефона, потому как все приложения устанавливаются только сюда, а и не на SD карту.
Но даже если места не хватает – постарайтесь его освободить, потому как именно обновления отвечают за корректную работу приложения
. Цель новых версий не только в том, чтобы придумать нововведения, но и исправить ошибки старых версий. После того, как приложение установило обновление, программой опять можно пользоваться.
После того, как приложение установило обновление, программой опять можно пользоваться.
Скорый редизайн мобильных приложений и iOS. Первые концепты обновлений внешнего вида опубликованы в официальном блоге «ВКонтакте», в котором, помимо всего прочего, Князев также рассказал о работе дизайнеров в компании.
Помимо кардинально нового пользовательского интерфейса, дизайнеры «ВКонтакте» с грядущим обновлением внешнего вида добавят и несколько функциональных нововведений. В разделе «Поиск и рекомендации» появятся прямоугольные истории пользователей, находящихся поблизости, а также лента потенциально интересных записей от новых авторов.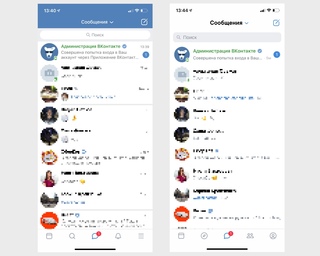
Когда выйдут обновленные
Они получили новый дизайн, где больше нет бокового меню, новый раздел с лентой рекомендаций, а в будущем компания собирается запустить платформу для создания лонгридов внутри соцсети. провёл сутки с новым приложением и пообщался с представителями компании о том, как теперь всё будет работать.
В веб-версии соцсети у ленты новостей уже был раздел «Рекомендации», но его контент не синхронизирован со вкладкой поиска в приложении, где работает новый алгоритм. Кроме того, контент во вкладке визуально группируется: фотографии и видео укладываются плиткой, записи с длинными текстами занимают больше пространства и выделяются.
Примерно по половине записей можно сразу сказать, как они попали в рекомендации — в основном из-за того, что их лайкнули или прокомментировали друзья. Другая половина попадает неведомым образом: мне алгоритм в топе рекомендаций показал несмешное (хотя вроде как юмористическое) видео неизвестной девушки с 43 просмотрами и одним лайком даже не от друга.
Другая половина попадает неведомым образом: мне алгоритм в топе рекомендаций показал несмешное (хотя вроде как юмористическое) видео неизвестной девушки с 43 просмотрами и одним лайком даже не от друга.
В ленте есть и множество коротких текстовых записей, иногда встречаются публикации знаменитостей с верифицированными профилями (у меня это были Руслан Соколовский и Илья Варламов). В целом рекомендации оставляют впечатление более удобного для потребления контента инструмента, чем лента новостей или видеораздел. За счёт убранных не обязательных в данном случае элементов интерфейса контент вышел вперёд и стал более опрятным. По нажатию на карточку можно перейти в саму запись — там будет доступен весь привычный интерфейс и кнопка для быстрой подписки.
«ВКонтакте» называет свой алгоритм «Прометеем» (в древнегреческих мифах он приносил людям огонь): когда пост пользователя попадает в рекомендации, он получает метку в виде пламени на своей странице. Так можно будет понять, откуда пошёл поток новых комментариев и лайков.
Но в ленте рекомендаций работает не только «Прометей», который ищет что-то новое и поддерживает малоизвестных блогеров. Есть ещё два типа подбираемого контента: первый — персональные рекомендации, построенные на социальном графе пользователя, а второй — общие для «ВКонтакте» тренды.
Представители «ВКонтакте» рассказали, что «Прометей» умеет искать пользователей, подобным другим. Если соцсеть знает, что у пользователей популярен художник Дюран, она будет использовать собранные об их интересах данные, чтобы искать похожих на него блогеров.
Вместе с тем компания обещает, что будет бороться с плагиаторами, которые не производят свой контент, а зарабатывают популярность на краже его у других. Алгоритм будет искать оригинал и стараться показывать именно его, причём речь идёт о всех медиа — тексте, картинках и видео, поясняет Сергей Паранько, развивающий медиаплатформу соцсети: «Если кто-то взял чужой контент и начал на нём подниматься, то мы всегда проверим — а не был ли этот контент загружен кем-то ранее».
Косметика
В начале сентября ведущий дизайнер «ВКонтакте» Павел Князев анонсировал скорый редизайн мобильных приложений. В текущем виде приложения реализовано только половина из обещанного: лайки стали красными (выглядят гораздо сочнее, нажимать приятнее), кнопки стали более круглыми, перерисовано много иконок и, главное, в приложении больше нет бокового меню — есть только пять кнопок снизу.
Визуально это непривычно: разделы бокового меню теперь находятся под пятой кнопкой, и туда же спрятали музыку и видео: при прослушивании музыки придётся возвращаться именно в эту вкладку, чтобы переключать треки. Дизайн аудиоплеера пока прежний, а в видеоразделе до сих пор нет рекомендаций, которые появились на десктопе ещё в 2015 году — теперь все рекомендации только во вкладке поиска.
Лента новостей очень напоминает комбинацию Фейсбука и Инстаграма: с одной стороны, здесь есть разнокалиберный «тяжёлый» контент, выдача которого находится под управлением алгоритма. С другой — истории и быстрый доступ к их созданию по свайпу вправо. Но теперь у «ВКонтакте» есть не одна, а уже две алгоритмических ленты в соседних вкладках: новости и рекомендации.
С другой — истории и быстрый доступ к их созданию по свайпу вправо. Но теперь у «ВКонтакте» есть не одна, а уже две алгоритмических ленты в соседних вкладках: новости и рекомендации.
Звучит непросто, тем более что полной синхронизации с десктопом мобильная версия не предполагает — ни в методах работы, ни в визуальном оформлении. Управляющий директор «ВКонтакте» Андрей Рогозов поясняет: веб давно ушёл на второй план, впереди — мобильные платформы, да и паттерны пользователей на них сильно различаются.
Платформа для лонгридов
Пока у «ВКонтакте» есть лишь старые возможности для создания записей — написание текстовых постов, съёмка фотографий, видео и историй с масками. Но до конца 2017 года соцсеть обещает запустить платформу для создания лонгридов, которые будут «корректно отображаться на всех устройствах».
Паранько пояснил, что платформа для лонгридов — это по сути редактор длинных записей с различными вложениями, который позволит публиковать внутри соцсети сложные форматы. Раньше паблики «ВКонтакте» использовали для этого wiki-разметку, но она, скорее всего, канет в прошлое — её не отключат, но развивать не будут. По описанию платформа будет напоминать то, что уже существует у Medium, Facebook и Telegram.
Раньше паблики «ВКонтакте» использовали для этого wiki-разметку, но она, скорее всего, канет в прошлое — её не отключат, но развивать не будут. По описанию платформа будет напоминать то, что уже существует у Medium, Facebook и Telegram.
У Medium это каталог интересных статей, у Facebook — инструмент более богатой вёрстки текста, а у Telegram — примитивный редактор для публикации любых больших материалов с поддержкой быстрой загрузки через Instant View. Во «ВКонтакте» заявляют, что будут поддерживать инициативу AMP (этот стандарт используют Google и Twitter) для более быстрой загрузки своих материалов, в том числе встроенных на внешние сайты.
По задумке компании, для пользователя чтение внешних материалов и опубликованных через платформу не должно ничем отличаться — причём «умная» лента будет ранжировать такие записи выше, чем обычные ссылки, так как для пользователя контент будет загружаться быстрее. По статистике «ВКонтакте», за год работы «умной» ленты на неё перешло 85% пользователей.
Насколько корректным будет отображение на всех устройствах — пока вопрос. Ещё несколько лет назад в рамках аналогичной инициативы для медиа во «ВКонтакте» появились большие карточки у ссылок на материалы СМИ и других внешних сайтов — однако приложение соцсети для iPad до сих пор не умеет их корректно отображать.
Встроенный мессенджер
Ещё в августе 2014 года представители «ВКонтакте» во время специального мероприятия заявили о планах выпустить отдельный мессенджер для обмена сообщениями. Эти планы многократно повторялись источниками в СМИ вплоть до конца 2016 года, пока вопрос «Когда “ВКонтакте“ выпустит мессенджер» окончательно не стал мемом.
Неформально представители соцсети уже давно признают, что необходимость в отдельном мессенджере отпала. Теперь это закреплено в официальных пресс-релизах компании: она стала отдельно считать аудиторию «встроенного мессенджера ВКонтакте» — по последним данным, это 82 миллиона человек в месяц. Ежедневно соцсетью пользуется 97 миллионов человек.
Как обновить ВК на компьютере до последней версии в 2020 году
Где можно скачать ВК QR код
Администрация ВКонтакте сообщает, что сегодня приходят сотни писем с просьбой объяснить как можно установить новую версию ВК. Также сообщается, что поступает большое количество жалоб о том, что новая версия ВКонтакте недоступна. Дело в том, что если просто зайти в Play Market и скачать приложение, то воспользоваться последними .обновлениями не получится. Обнова еще находится в состоянии бета-тестирования и недоступна рядовым пользователям.
Как бы там ни было, посмотреть обновление можно, если воспользоваться одной из следующих инструкций:
Инструкция без QR кода
- В первую очередь требуется установить последнюю версию приложения с Плей Маркета. Для системы Андроид: «ВКонтакте — общение, музыка и видео«
- Для iOS: «VK — общение, музыка и видео«
 up/future
up/futureИнструкция по обновлению при помощи QR кода
Обновить через QR очень просто. Нужно просто навести телефон на указанный ниже код, после чего, приложение автоматически начнет обновление. Как это сделать?
Следуйте инструкции:
- Как и в предыдущий раз — устанавливаем последнюю версию приложения с Play Market: Android: «ВКонтакте — общение, музыка и видео«
- iOS:«VK — общение, музыка и видео«
 Пользуйтесь.
Пользуйтесь.Дополнительная информация
Любое обновление, которое выходит в соц.сети, предоставляется бесплатно для пользователей. Часто нововведения сложно даются для изучения, многие не сразу могут привыкнуть к измененному интерфейсу, из-за чего разработчики оставили функцию возврата.
Если не понравилась обнова, то вернитесь на старую версию, нажав на ссылку в том же месте. Отказаться можно на любом устройстве. Но, когда программисты доработают все недочеты и убедятся в стабильности, все пользователи принудительно будут переведены на новую версию.
Как выглядит редизайн ВК после обновления 2020
Также, многие люди задаются вопросом — как будет выглядеть новая версия обновленного ВКонтакте, так как возникает вопрос «А стоит ли вообще заморачиваться?» Обновленный ВК выглядит следующим образом:
Стоит также отметить, что обновленная версия позволяет вернуться к предыдущему оформлению. Так что в случае чего всегда можно вернуть предыдущий вид страницы.
Сбой браузера
Почему постоянно обновляется страница «ВКонтакте»? Следующая причина — это сбой браузера. Не стоит путать данный вариант с обычным системным сбоем. Ведь в действительности между данными понятиями прослеживается серьезная разница.
Впрочем, если говорить о путях исправления ситуации, то методы при системном сбое и браузерном тоже отличаются. Именно поэтому данные варианты придется научиться различать. Для пользователя они не столь существенны. Определить, какой именно сбой имеет место, бывает весьма проблематично. Поэтому сначала стоит исключить браузерный, а затем — системный. Как это сделать? О способах борьбы с проблемой — немного позже.
Во время обновления ВКонтакте через QR код произошла ошибка — что делать?
Если во время установки на устройстве произошла ошибка и программа не обновилась необходимо совершить действия, описанные ниже для устранения неполадки.
- Проверьте все ли действия совершены как указано в инструкции. Установлена ли последняя версия приложения перед обновлением
- В предыдущие года также было много обновлений ВК, в связи с чем убедитесь, что использован верный QR код 2020
- Просканируйте устройство на наличие вирусов
- Очистите от «мусора» телефон, удалите ненужные программы
- Проверьте скорость и наличие интернет-соединения
тематические ленты, дизлайки, донаты и многое другое
Сегодня на мероприятии VK Content Day социальная сеть анонсировала ряд крупных обновлений, среди которых запуск тематической ленты с отбором контента экспертами, дизлайки в комментариях новые возможности для монетизации, публикация историй для всех сообществ и многое другое.
«Два года назад мы начали активно работать с создателями контента — и с тех пор стремительно развиваемся в этом направлении. Фундаментом для роста контентной платформы ВКонтакте станут новые возможности, которые мы представили на VK Content Day, и обновлённое приложение. Мы продолжим создавать для авторов и медиа нужные им инструменты, помогать находить аудиторию и зарабатывать, а пользователям — получать интересный тематический контент», — Андрей Рогозов, CEO «ВКонтакте».
Тематические ленты и открытая редакция
ВКонтакте представила тематические ленты — это вкладки в «Рекомендациях», где собираются записи по конкретным темам: играм, IT, туризму, спорту, юмору, стилю и другим. Чтобы в разделе появлялось всё самое интересное, мы объединили усилия людей и алгоритмов.
Тематические ленты курируют участники открытой редакции. При этом у каждой темы свой состав экспертов. Чтобы стать одним из них и влиять на рекомендации, достаточно быть активным пользователем ВКонтакте и глубоко знать какую-то область.
Тематику публикаций определяют алгоритмы, но автор может указать её и самостоятельно при создании или редактировании записи. Эксперты голосуют за или против материалов по своей теме. Чем больше положительных откликов от участников редакции, тем выше алгоритмы поднимут публикацию в «Рекомендациях».
Если эксперт неактивен или на материалы, которые он поддержал, поступает много жалоб, участника могут исключить из редакции. О том, как стать одним из экспертов, можно будет прочесть в сообществе ВКонтакте с авторами, когда откроется набор.
Также в качестве эксперимента ВКонтакте представит собственное издание — «Фокус». Оно будет доступно в виде паблика и вкладки в разделе «Интересное» в новой версии мобильного приложения VK.
Увеличение жизни качественных записей
ВКонтакте пересмотрит один из принципов умной ленты — в первую очередь предлагать свежие публикации. Ведь по-настоящему качественный контент остаётся интересным и спустя время. Поэтому, если материалы высоко оценит «ядро аудитории», они будут показываться в лентах новостей и рекомендаций наравне с недавно созданными записями — и смогут собирать охваты гораздо дольше.
Дизлайки в комментариях
Подписчики смогут выразить своё несогласие с мнением комментатора, но не вступать при этом в дискуссию — просто поставить дизлайк.
Новый способ монетизации — VK Donut
С помощью VK Donut подписчики смогут финансово поддерживать любимых авторов и интересные медиапроекты. Ежемесячная подписка оформляется прямо ВКонтакте.
В благодарность авторы могут предоставлять пользователям с платной подпиской на сообщество эксклюзивные возможности. Например, открывать им ранний доступ к записям и статьям, проводить розыгрыши призов, встречи или камерные концерты, публиковать специальные материалы. Представители бизнеса могут делать закрытые распродажи и акции, а также организовать программу лояльности именно для клиентов с подпиской.
Бонусы для поклонников и минимальный размер доната администраторы сообществ настраивают самостоятельно. Получать деньги можно на счёт VK Pay и на банковские карты. На старте комиссия будет самой низкой на рынке — 5%, однако в будущем она может быть пересмотрена.
Верификация 2.0
ВКонтакте изменит подход к верификации. Раньше галочка была символом особой популярности — небольшие заведения или локально известные артисты не могли её получить. Теперь отметка верификации станет только подтверждением, что страница официальная: её ведут настоящие представители, а не фанаты или злоумышленники. Так станет проще найти сообщество любимой уютной кофейни или подписаться на многообещающего, но пока начинающего музыканта.
Пройти верификацию можно будет прямо в настройках профиля или сообщества — достаточно подтвердить известность на локальном уровне, и закрепить связь со страницей с помощью документов.
Обновление приглашений на мероприятия
Чтобы пользователи получали приглашения именно на мероприятия, которые им интересны, ВКонтакте начнёт выдавать их сообществам частями. Организатор сможет рассылать небольшое количество приглашений и получит следующую партию, только если получатели будут реагировать позитивно: принимать их, переходить в сообщество, не жаловаться на спам. Так порядочные администраторы ничего не потеряют, а благодаря более точным рекомендациям аудитория будет относиться к приглашениям с большим доверием.
Так порядочные администраторы ничего не потеряют, а благодаря более точным рекомендациям аудитория будет относиться к приглашениям с большим доверием.
Истории для всех сообществ и создание интерактивных стикеров в мини-приложениях
Уже в этом году все сообщества смогут публиковать истории. Однако они будут сгруппированы — так подписчики не потеряются в обилии контента. Интересные страницы пользователь сможет перенести в основной блок, а два сообщества, где подписчик наиболее активен, закрепятся автоматически.
Группироваться будут и одинаковые истории, созданные в сервисах VK Mini Apps. Кроме того, с помощью сервисов можно будет создавать новые интерактивные стикеры и прикреплять их к историям. Если пользователь нажмёт на такой стикер, то сможет, не отрываясь от истории, записаться на приём к врачу, вступить в беседу, оставить комментарий или купить товар. iGuides в Telegram — t.me/igmedia iGuides в Яндекс.Дзен — zen.yandex.ru/iguides.ru У нас есть подкаст и его видео-версия
www. iguides.ru
iguides.ru
Промо посты — новый вид рекламы в ВК: обьзор
Уже как месяц назад с социальной сети «ВКонтакте» начал свою работу новый вид рекламных объявлений – «промопосты». Это так называемые промоуторские посты, по сути, обычные рекламные объявления, но представлены они в новом формате.Уже как месяц назад с социальной сети «ВКонтакте» начал свою работу новый вид рекламных объявлений – «промопосты». Это так называемые промоуторские посты, по сути, обычные рекламные объявления, но представлены они в новом формате. Генеральный директор социальной сети Борис Добродеев, пояснил, что теперь рекламные объявления можно размещать прямо в новостной ленте пользователя. Такой вид рекламы давно используется в Twitter и Facebook.
Данное нововведение не стало неожиданностью. О введении подобного вида рекламы сообщалось на конференции «РИФ+КИБ 2015» в апреле этого года. Глава отдела по работе с клиентами «ВКонтакте» Александр Круглов пояснил, что данный вид рекламы не будет перегружать новостную ленту пользователя, так как введено ограничение на количество показанных промопостов. Так, первый промопост появится в ленте новостей через 5-10 сообщений, последующие, через каждые 25. В отличие от других социальных сетей, такое ограничение сделает рекламу менее навязчивой.
Так, первый промопост появится в ленте новостей через 5-10 сообщений, последующие, через каждые 25. В отличие от других социальных сетей, такое ограничение сделает рекламу менее навязчивой.
Введение промопостов станет отличным подарком для рекламодателей, которые смогут размещать рекламные посты на легальной основе без участия администраторов тематических сообществ.
В чем суть промопостов?
Промопост – это разновидность, так называемой «нативной» рекламы. Данный формат подает рекламные сообщение как естественную часть сайта. Все чаще, рекламные посты выглядят не как рекламные сообщения, а как мнения или советы от обычных пользователей, что вызывает больше доверия к рекламе в целом.
Более того, в промопостах есть все возможности обычных постов. Вы можете размещать в них текст, аудио-видео контент, фотографии, карты и т.д.
На сегодняшний день промопосты доступны только для компьютерной версии «ВКонтакте». Разработчики уверяют, что совсем скоро промопосты появятся и в мобильных версиях. Это позволит сделать промопосты кроссплатформенными.
Это позволит сделать промопосты кроссплатформенными.
Главные особенности промопостов
В чем же основные различия промопостов от обычных?
- Промопост можно будет лайкнуть, но нельзя комментировать.
- Промопост можно будет показывать для определенной категории пользователей. Т.е. промопост можно будет размещать для определенной аудитории по множеству критериев.
- Рекламодатели, которые воспользуются промопостами, получат доступ к 20 видам таргетинга. Что существенно увеличит эффективность данного вида рекламы.
- Промопосты – платная услуга. Оплата такой рекламы производится за каждые 1000 пользователей, которые просмотрят рекламное сообщение по модели uCPM. Так же планируется использование других моделей: плата за переходы или плата за просмотр видеороликов и тд.
Новый вид мошенничества в ВК. Как меня сегодня развести на деньги пытались | Мама — Манчижужа🐞
Всем привет, на связи Мама-Манчижужа 🐞, и сегодня я Вам расскажу занятную историю. 😊
😊
Каждый из нас состоит в какой-то виртуальной секте 🗿 : кто в ВК (Вконтакте), кто в ОК (одноклассники), а еще есть Фейсбуки, инстаграмы, Тик-токи…
Да даже Дзен — та же секта! 🙏 😄
Как попадешь сюда, так и залипнешь на час, а появится свободная минутка — оп, уже снова ленту листаешь…
В онлайне у меня два «греха»: Дзен и ВК. На остальные соцсети у меня просто нет времени!
Не понимаю, как люди успевают сидеть на пяти своих страничках, выкладывать фото, читать про жизнь чужих им людей? При чем заходят туда ежедневно! КАК, ну КАК Вы это делаете?? 🙉
Сегодняшний пост посвящен ВК, а точнее мошенничеству, которое можно провернуть с его помощью.Каждый из Вас подписан на десяток, а то и полтинник интересных сообществ. И вот, некоторые группы время от времени устраивают розыгрыши подарков. Честно признаться, я люблю подарки на халяву.
И мне даже удавалось выигрывать!
Как я выиграла набор органической косметики можно прочитать ЗДЕСЬ
Но всегда нужно быть внимательным и не кидаться на все розыгрыши подряд!
Я давно мечтаю о швейной машинке, компактной такой, современной. и тут мне приходит сообщение от подруги: мол смотри, как ты и хотела машинку разыгрывают! Итоги уже сегодня, успевай!
и тут мне приходит сообщение от подруги: мол смотри, как ты и хотела машинку разыгрывают! Итоги уже сегодня, успевай!
Конечно, я решила испытать удачу, тем более участников было очень мало, что-то около ста. Обычно за такие большие призы «борются» несколько тысяч человек. А тут 100, тьфу, да я уже почти шью на этой машинке, настолько велик шанс на победу! 😂
Выполнила условия, пробежалась по группе: много, ооочень много постов выкладывается в течение дня. Обычно так не ведут сообщества. Это был первый звоночек, что где-то тут подвох))
На следующий день мне приходит сообщение, типа от админа группы, который сообщает мне о победе!
Да ладно, серьезно?? Ехху!!!
Но я решила удостовериться в этом, и перешла на страницу группы. Там я не увидела поста об итогах, и это снова посеяло во мне сомнения. А еще не было и самого поста с розыгрышем…задала прямой вопрос)
Получила ответ, правда заскринила немного коряво, но все понятно должно быть))
ВАУ, ЭТО ВСЕ МОЕ!!! 😄
Я уже на 99% была уверена, что это мошенники! Решила еще их немножко попытать и сказать, что у меня есть кому забрать из Сегежи МОЮ машинку!
После этого мошенники решили, что с меня им поиметь ничего не удастся и внесли в черный список! 😄
Как так? Я же победитель! Разве так поступают с победителями? 😁
Вобщем, написала в техподдержку ВК, чтоб проверили эту группу. Я подозреваю, что они так всем участникам отписались. Собрали личные данные для каких-то целей, а может еще и денег за пересылку приза попросили. Кто-то да отправит, я уверена в этом!
Я подозреваю, что они так всем участникам отписались. Собрали личные данные для каких-то целей, а может еще и денег за пересылку приза попросили. Кто-то да отправит, я уверена в этом!
Честные группы и розыгрыши существуют, но и нарваться на мошенников в сети легко!
Будьте внимательны, всегда перепроверяйте информацию и никогда не отправляйте деньги за свой честно выигранный приз! 😉
Как переключиться на старый дизайн вк. Как изменить новый дизайн вк на старый за считанные секунды
Доброго всем времени суток, мои дорогие друзья. С вами как обычно Дмитрий Костин. И сегодня я хочу снова поговорить про контакт, то есть про социальную сеть «Вконтакте». Не так давно (в августе 2016 года), как вы знаете, абсолютно у всех сменился дизайн вк. Это очень взбудоражило весь рунет. Кому-то новый вид нравился, но большинство он раздражал и многие просто плевались, глядя на него и задавались вопросом: «А можно ли вернуть старую версию?».
Мне, если честно параллельно на всё это. И на самом деле я ничего плохого в нем не вижу. По-моему смотрится вполне неплохо, несмотря на то, что новый вид у большинства людей ассоциируется со смесью одноклассников и фейсбука. Ну тут конечно доля правды есть). Но я не об этом.
В общем, если всё таки новый стиль вам не нравится, если он мозолит вам глаза, то я могу вам рассказать, как изменить дизайн вк на старый за считанные секунды. Да, вы можете это сделать. Конечно — это не официальная фишка, но я думаю, что вас это особо волновать не будет. Всё, что нам понадобится — это установить специальное расширение в браузер. Я буду показывать всё на примере гугл хрома. Итак, поехали!
Первый вариант
Ну ка, зайдите теперь в свой аккаунт контакта и посмотрите. Ну как? Нормально. Вроде похоже, хотя есть во многих местах неточности и косячки, но они не так бросаются в глаза. Так что можете восстановить прежнюю версию, если вам старый вид роднее и удобнее.
Второй вариант
Этот способ также предусматривает установку расширения, но уже другого. И чтобы его активировать потребуется чуть больше времени. Я не буду описывать процесс установки самого расширения, так как всё это написано выше. В общем поехали!
И чтобы его активировать потребуется чуть больше времени. Я не буду описывать процесс установки самого расширения, так как всё это написано выше. В общем поехали!
Вы можете включить разные виды и переключаться между ними с помощью самого расширения stylish. Тут уже как сами пожелаете.
Кстати это расширение (дополнение) присутствует и в других браузерах, так что если вы пользуетесь например Mozilla Firefox или Оперой, то вам не составит труда поставить всё то же самое.
Если у вас остались какие-то непонятки, то вы можете посмотреть мою видеоинструкцию.
Ну как вам? По-моему очень круты расширения, и ставятся они за считанные секунды, а если он вам больше не понадобятся, то можете их просто отключить). Какое из-них вам понравилось больше? Или может у вас есть свой способ? Поделитесь своим мнением в комментариях.
Ну а на этом я свою статью пожалуй буду заканчивать. Надеюсь, что она была для вас полезна и интересна. Кстати не забываем подписываться на обновления моего блога, чтобы всегда быть в курсе всех важных событий и интересных статей на блоге. Увидимся с вами в других статьях. Пока-пока!
Увидимся с вами в других статьях. Пока-пока!
С уважением, Дмитрий Костин
После длительного тестирования новый дизайн «ВКонтакте» был применен для всех пользователей социальной сети. Понравилось кардинальное изменение внешнего вида «ВКонтакте» далеко не всем, при этом стандартного средства для возвращения на старый дизайн разработчики не предусмотрели, что вызвало бурю негодования среди пользователей. К счастью, существует способ, который позволяет вернуть привычный дизайн социальной сети. О нем мы в этой инструкции и расскажем.
Перед тем как начать, хотим обратить ваше внимание на то, что старый дизайн «ВКонтакте» возвращается частично. Происходит так из-за того, что стиль старого внешнего вида на текущий момент до конца недоработан. Тем не менее, это единственный рабочий способ возвращения старого дизайна «ВКонтакте» — выбирать не приходится.
Шаг 1. Загрузите и установите расширение Stylish для Chrome или Firefox .
Шаг 2. Установите стиль старого дизайна «ВКонтакте», перейдя по этой ссылке .
Шаг 3. Откройте «ВКонтакте» или перезагрузите уже запущенную страницу социальной сети.
Изменения вы увидите сразу — установленный стиль вернет старый дизайн социальной сети практически полностью! Все недоработки разработчики стиля в ближайшее время исправят и вам нужно будет только лишь обновить стиль при помощи расширения Stylish.
Как обновить стиль старого дизайна «ВКонтакте»
Шаг 1. Нажмите на иконку расширения Stylish.
Шаг 2. Выберите пункт «Управление установленными стилями »
Шаг 3. Нажмите кнопку «Проверить обновления ».
Смотрите также:
Поставьте оценку:
В апреле 2016 года началось тестирование нового дизайна ВК. Первое время пользователи могли отказаться от него, продолжив взаимодействовать со старым интерфейсом. Но сейчас почти все странички отображаются в новом виде, поэтому у многих возник вопрос, как вернуть старый дизайн ВКонтакте.
В приложениях изменить дизайн нельзя, можно пользоваться только тем интерфейсом, который предложили разработчики.
В браузерной версии раньше можно было откатиться к привычному дизайну одним нажатием кнопки или удалением приставки «new» из адреса сайта.
Сейчас эти способы не работают, хотя старая версия странички никуда не пропала. Не помогает и замена приставки «new» на «0» или переключение на мобильную версию сайта – интерфейс остается новым. Но есть всё-таки несколько проверенных способов, которые помогут еще на время оставить привычный интерфейс.
Ссылка на страницу
Самый простой способ открыть свою страничку в старом оформлении – перейти по ссылке http://philka.ru/vk.php . В результате перенаправления откроется ваш профиль, но вместо нового «фейсбучного» интерфейса вы увидите привычный интерфейс, который не менялся серьезно очень давно.
Главный недостаток этого метода – ограничение его действия на одну сессию в браузере.
Проще говоря, как только вы закроете веб-обозреватель, внесенные изменения пропадут, и при следующем входе на vk. com откроется новая версия сайта.
com откроется новая версия сайта.
Для удобства можете добавить эту инструкцию в закладки и постоянно переходить по ссылке. Чтобы сделать закладку, нажмите на звездочку в адресной строке, а затем подтвердите сохранение странички.
Ярлык для запуска
Если вариант с закладкой не устраивает, создайте отдельный ярлык, при открытии которого будет запускаться браузер с вашей страничкой ВК в старом оформлении.
Чтобы открыть профиль ВК со старым интерфейсом, щелкните дважды по созданному ярлыку. Запустится браузер (или откроется новая вкладка, если браузер уже был запущен), в окне которого вы увидите привычный дизайн.
Скрипт старого дизайна
Если вы не хотите постоянно искать ссылку на старый ВК или создавать ярлыки со скриптами, запускающими браузер с нужной страницей, то воспользуйтесь способом, который стал доступен благодаря светлой голове Николая Исаева. Он создал специальный скрипт «Старый Дизайн ВКонтакте», который можно установить в браузере.
После добавления скрипта сайт будет всегда открываться с прежним интерфейсом, не нужно будет никаких ссылок и дополнительных ярлыков.
Но чтобы скрипт начал работать, необходимо предварительно добавить в браузер возможность выполнения пользовательских скриптов. По умолчанию эта функция недоступна, но это можно исправить с помощью специальных расширений.
- Opera, Яндекс.Браузер – Violent monkey.
- Google Chrome, Яндекс.Браузер, Safari – Tampermonkey.
- Mozilla Firefox – Greasemonkey.
Найти расширение можно в магазине приложения браузера. Распространяются все эти дополнения бесплатно, так что возврат старого интерфейса ничего не будет вам стоить. После добавления расширения для установки пользовательских скриптов перейдите на страничку «Старый Дизайн ВКонтакте» и нажмите «Установить этот скрипт».
После успешной установки при переходе на сайт vk.com будет открываться старая версия странички. Вы можете закрывать браузер, перезагружать компьютер, ничего не пропадет. Скрипт нужно обновлять, потому что периодически автор вносит в него изменения и исправления. Апгрейд делается через расширение, которое вы установили перед добавлением скрипта.
Скрипт нужно обновлять, потому что периодически автор вносит в него изменения и исправления. Апгрейд делается через расширение, которое вы установили перед добавлением скрипта.
Чтобы удалить скрипт, перейдите в панель расширения. В таблице установленных скриптов найдите «Старый Дизайн ВКонтакте» и нажмите «Удалить».
Можно пойти более радикальным путем и удалить не скрипт, а само расширение. Скрипт в таком случае перестанет работать, а ВКонтакте будет открываться с новым дизайном.
ВК новый дизайн — ВКонтакте представила новую версию приложения для смартфонов
Логотип ВКонтакте / ВКонтактеКоманда ВКонтакте (VKontakte) официально представила новый дизайн ВК в приложении для смартфонов. У части пользователей он уже доступен, массовый релиз состоится в течение недели.
У ВКонтакте обновился дизайн абсолютно всех разделов. Теперь приложение ВК для iOS и Android выглядит более современно, а также стало удобнее и прибавило в возможностях.
Основное отличие — измененная нижняя панель. Там теперь расположились пять новых вкладок: «Новости», «Обзор», «Сообщения», «Друзья» и «Профиль».
Там теперь расположились пять новых вкладок: «Новости», «Обзор», «Сообщения», «Друзья» и «Профиль».
В разделе «Новости» можно найти ВК сторис, посты из различных групп и пабликов, мемы, видео, а также вкладку «Интересное», которая заменила ленту рекомендаций.
Внешний вид вкладки «Новости» / ВКонтактеВкладка «Обзор» пригодится для различных сторонних сервисов, связанных с ВК. ВКонтакте новый дизайн объединил все приложения в одном месте. Именно тут вы сможете слушать музыку ВК, смотреть трансляции, пересылать деньги и выполнять множество других задач.
Внешний вид вкладки «Обзор» / ВКонтактеС вкладкой «Сообщения» все предельно ясно. Именно тут вы будете общаться с друзьями ВКонтакте. Дизайн мессенджера изменился, чтобы общение было более удобным.
Сравнение старого дизайна сообщения (слева) и нового (справа) / ВКонтактеВкладка «Друзья» теперь тоже вынесена на основную панель, подчеркивая важность товарищей ВКонтакте. Именно там вы найдете всех ваших знакомых.
Внешний вид вкладки «Друзья» / ВКонтактеИ самая последняя вкладка на нижней панели — это «Профиль», в котором указана вся информация о вас. Теперь добавлять друзей можно просто через QR-код в вашем профиле.
Теперь добавлять друзей можно просто через QR-код в вашем профиле.
Все недостающие возможности переехали в боковое меню. Оно доступно через «Профиль». Там надо просто смахнуть справа налево.
Боковое меню / ВКонтактеЕсли вы задаетесь вопросом, как вернуть старый ВК, то ответ вряд ли вас обрадует. Именно новая версия теперь считается стандартной, так что все обновления безопасности и нововведения будут только в ней. На iOS откатиться почти невозможно, а на Android необходимо найти специальные apk-файлы, в которых и будет ВК старая версия.
Ранее мы рассказывали, как скачать музыку и видео с ВКонтакте.
«Драгоценный» новый вид, названный в честь Голлума Властелина Кольца
Рыба была найдена в Керале, Южная Индия, и является первой змееголовой рыбой, которая, как было описано, живет под землей, ее совместное подземное существование с Голлумом, как сообщается, быть причиной его наименования.
Змееголовы происходят из Африки и Азии и отличаются длинными спинными плавниками, большими ртами и способностью дышать воздухом, используя так называемый наджаберный орган в их жаберной полости.
Эти рыбы обычно живут в пресноводных реках или водно-болотных угодьях, что делает это новое открытие особенно захватывающим.
Этот вид был обнаружен случайно после того, как фотографии рыбы были размещены в Интернете после разрушительного наводнения, обрушившегося на Кералу в августе прошлого года.
Эти изображения побудили Керальский университет рыболовства и океанологических исследований провести расследование.
«Как только я увидел изображения более высокого качества, стало ясно, что это змееголов», — говорит доктор Ральф Бритц из Музея естественной истории и соавтор статьи, описывающей новый вид.
«Змееголов из подземной среды обитания — захватывающая находка. Эта рыба очень необычная ».
Aenigmachanna gollum. © Ануп В.К.
В глобальном масштабе подземные рыбы в основном представлены только двумя группами: сомами и карпами, поскольку виды в этих группах считаются предварительно приспособленными к жизни под землей.
Змееголовы, однако, не склонны иметь какие-либо приспособления, которые также подходили бы для подпольного образа жизни.
«Удивительно то, что — если предположить, что это подземная рыба — у нее нет типичных адаптаций, которые вы ожидаете от подземной рыбы», — объясняет Бритц.
«Либо рыба только недавно приобрела этот подземный образ жизни, либо она живет в какой-то переходной зоне, где все еще перемещается между подземной и наземной средами обитания. Мы просто не знаем, поэтому нам было бы очень интересно узнать об этом ».
Новая рыба очень самобытна по сравнению с известными змееголовыми, у нее невероятно длинное, напоминающее угря тело; многочисленные чешуи, очень длинный анальный плавник, идущий вдоль брюха и хвоста, утративший способность сохранять плавучесть в толще воды.
В настоящее время это восьмой вид подземных рыб, вышедших из водоносных горизонтов под Кералой, что заставляет исследователей полагать, что должна существовать скрытая экосистема, существующая на глубине более 40 метров под поверхностью.
В августовском выпуске журнала BBC Wildlife Magazine за август 2018 года также освещалось открытие нового вида, названного в честь персонажа «Властелин колец» ; Креветки Бильбо Бэггинса, названные в честь сходства с хоббитом, у которого обе общие волосатые лапы, а в апреле 2017 года журнал BBC Wildlife Magazine сообщил об открытии первой пещерной рыбы в Европе.
Прочтите статью Zootaxa.
Основное изображение: Aenigmachanna gollum. © Ануп В.К.
Лягушки как переносчики Salmonella и Edwardsiella
Бхат П., Майерс Р. М. и Карпентер К. П. 1967. Edwardsiella tarda в исследовании диареи у подростков. — Дж. Hyg. (Камб.) 65 : 293–298.
Google Scholar
Эдвардс, П. Р. и Юинг, В.H. 1962. Идентификация Enterobacteriaceae. — Burgess Publ. Co., Миннесота.
Google Scholar
Ewing, W.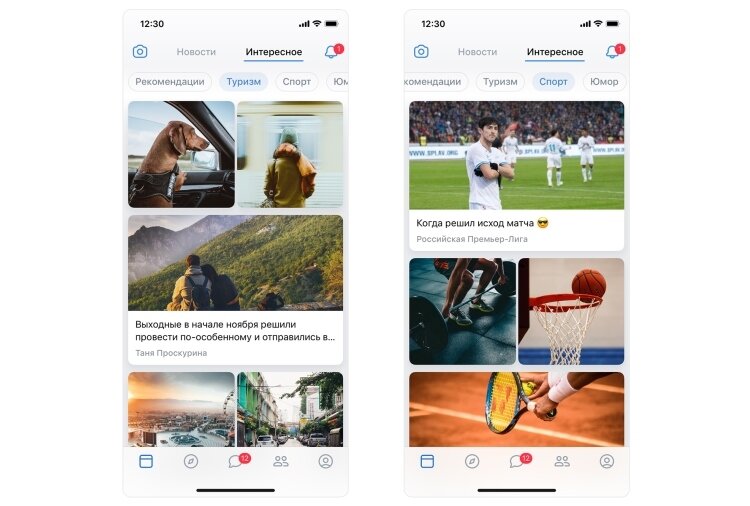 H., McWhorter, A.C., Escobar, M.R. и Lubin, A.H. 1965. Edwardsiella , новый род Enterobacteriaceae, основанный на новом виде, E. tarda . — Int. Бык. Бакт. Номенклатурный. Таксон. 15 : 33–38.
H., McWhorter, A.C., Escobar, M.R. и Lubin, A.H. 1965. Edwardsiella , новый род Enterobacteriaceae, основанный на новом виде, E. tarda . — Int. Бык. Бакт. Номенклатурный. Таксон. 15 : 33–38.
Google Scholar
Хеммес, Дж.H. 1966. Среда «7 дюймов», простая среда для быстрой предполагаемой идентификации как Salmonella , так и Shigella . — Антони ван Левенгук 32 : 442–446.
Google Scholar
Kauffmann, F. 1966. Бактериология Enterobacteriaceae. — Williams and Wilkins Co., Балтимор.
Google Scholar
Каура, Ю.К., Шарма, В.К. и Сингх И. П. 1970. Dabb-lizard Хардвика Uromastix hardwicki как резервуар Salmonella в природе. — Рез. Вет. Sci. 11 : 390–392.
Google Scholar
Каура, Ю. К., Шарма, В. К., Сингх, И. П., Саказаки, Р., Роде, Р. 1972. Змеи как резервуары Аризоны и Salmonella . — Zentralbl. Бактериол. Паразитенк. Инфектионскр. Hyg., I. Abt., Orig. 219 : 506–513.
Google Scholar
Саказаки Р., Тамура К., Прескотт Л. М., Бенчич З., Саньял С. К. и Синха Р. 1971. Бактериологическое исследование диарейного стула в Калькутте. — Indian J. Med. Res. 59 : 1025–1034.
Google Scholar
Шарма В. К., Каура Ю. К. и Сингх И. П., 1970. Аризона, инфекция у змей, крыс и человека. — Индиан Дж.Med. Res. 58 : 409–412.
Google Scholar
Шарма В. К., Каура Ю. К. и Сингх И. П. 1973. Дальнейшие наблюдения за статусом резервуара Salmonella ящерицы Dabb Хардвика. — Indian J. Med. Res. 61 : 184–189.
Google Scholar
Сонненвирт, А. С. 1970. Бактериемия с менингитом и без него, вызванная Yersinia enterocolitica, Edwardsiella tarda, Comamonas terrigena и Pseudomonas maltophilia .-Аня. Акад. Sci. 174 : 488–502.
Google Scholar
Sonnenwirth, A.C, и Kallus, B.A., 1968. Менингит, вызванный Edwardsiella tarda. — Amer. J. Clin. Патол. 49 : 92–95.
Google Scholar
Такал, Дж. В. Младший. и Менез, К. Ф. 1968. Выделение Edwardsiella tarda от собаки (Филипп. J. Vet. Med. 7 : 143–145.
Google Scholar
Уайт, Ф. Х., Нил, Ф. К., Симпсон, К. Ф. и Уолш, А. Ф. 1969. Выделение Edwardsiella tarda от страуса и австралийского сцинка. — Дж. Амер. Вет. Med. Жопа. 155 : 1057–1058.
Google Scholar
Dario neela, новый вид бадид-рыб из Западных Гат Индии (Teleostei: Percomorpha: Badidae)
Dario neela, новый вид бадид-рыб из Западных Гат Индии (Teleostei: Percomorpha: Badidae)Сожалеем, но GBIF не работает должным образом без включенного JavaScript.
Наш веб-сайт обнаружил, что вы используете устаревший небезопасный браузер, который не позволяет вам использовать этот сайт. Мы предлагаем вам перейти на современный браузер.
Домашняя страница набора данных
Цитата
Бритц Р., Ануп В. К., Даханукар Н., плази (2018).Дарио Нила, новый вид бадид-рыб из Западных Гат Индии (Teleostei: Percomorpha: Badidae). База данных таксономических обработок Plazi.org. Набор данных контрольного списка https://doi.org/10.11646/zootaxa.4429.1.6 доступен через GBIF.org 2021-04-19.Описание
Этот набор данных содержит оцифрованные обработки в Plazi на основе исходной статьи журнала Britz, Ralf, Anoop, VK, Dahanukar, Neelesh (2018): Dario neela, новый вид бадид-рыб из Западных Гат Индии (Teleostei: Percomorpha: Badidae ).Zootaxa 4429 (1): 141-148, DOI: https://doi.org/10.11646/zootaxa.4429.1.6Таксономические покрытия
Географические зоны покрытия
Библиографические ссылки
Контакты
Ральф Бритцавтор
В.K. Anoop
автор
Neelesh Dahanukar
автор
Гвидо Сауттер
Контактное лицо по административным вопросам
электронная почта: [email protected]
домашняя страница: http://plazi.org
издательство
Magnolia Press
St.Lukes 1346
Auckland
NZ
электронная почта: [email protected]
домашняя страница: http://www.mapress.com/
дистрибьютор
Plazi
Bern
CH
электронная почта: [email protected]
домашняя страница: http://plazi.org/
plazi
автор метаданных
Plazi
Aenigmachannidae, новое семейство змееголовых рыб (Teleostei: Channoidei) из подземных вод Южной Индии
Таксономия
Aenigmachannidae (змееголовые рыбы Gollum), новое семейство; Типовой род Aenigmachanna Britz, Anoop, Dahanukar & Raghavan 2019 (рис.1, 2).
Рисунок 1Aenigmachanna gollum . ( a ) Живая особь диаметром 60,6 мм (KUFOS 2019.8.226), зеленая стрелка отмечает положение вентиляционного отверстия; ( b ) КТ-изображение скелета голотипа 90,2 мм (BNHS FWF 966), зеленая стрелка отмечает положение вентиляционного отверстия, а красная стрелка — первый хвостовой позвонок; ( c ) КТ-изображение головы голотипа, желтыми стрелками отмечены заглазничный отросток и задний конец сильно удлиненной верхней челюсти; ( d ) КТ-изображение каудального скелета голотипа, обратите внимание на отсутствие кости Дей и наличие дистально раздвоенного гемального шипа (желтая стрелка) во втором предуральном центре; ( e ) КТ-изображение, окрашенное йодом 124.Образец 5 мм (KUFOS 2019.8.225) сбоку, плавательный пузырь показан синим цветом над пищеварительной системой (зеленый), обратите внимание на плавательный пузырь, заканчивающийся на уровне 8-го постанального позвонка.
Рис. 2Анатомия головы канала ( Parachanna , слева) и Aenigmachanna (справа) в сравнении. Нейрокраниум Parachanna africana , (MTD-F39824, c & s, нос не удален) 150 мм ( a , c , e ) и Aenigmachanna gollum , (KUFOS 2019.8,224) 81,8 мм в дорсальном ( a , b ), боковом ( c , d ) и вентральном ( e , f ) виде. Parachanna ( a ) с широким контактом между лобной и аутосфенозной, но Aenigmachanna ( b ) с широким контактом лобной и птеротической, за исключением аутосфеноза от крыши черепа. Parachanna с широкой крышей, образующей череп (отмечен белыми звездами в ( c ) над наджаберной полостью, саккулит показан фиолетовым, и простино-базиокципитальный шов слуховой капсулы отмечен фиолетовой стрелкой.Обратите внимание на наличие большого парасфеноидного зубного пятна и полной межглазничной перегородки, образованных контактом лобной части и парасфеноида у Aenigmachanna ( f ) и наличие межглазничного отверстия (отмеченного синим *) у Parachanna ( e ). Поперечные сечения КТ через голову Parachanna obscura , (CUMV 96491), длина неизвестна ( г ) и Aenigmachanna , 124,5 мм (KUFOS 2019.8.224), [вертикальные линии в ( g , h ) показан уровень разреза], чтобы проиллюстрировать наличие [отмечено черными звездами в ( g )] или отсутствие большой наджаберной полости.Виртуальные слепки внутреннего пространства рта и жаберной полости подтверждают наличие крупных парных наджаберных полостей у Parachanna ( i ) и их отсутствие у Aenigmachanna ( j ).
Диагноз: Семейство акантоморфной клады Labyrinthici (Anabantiformes), о чем свидетельствует общее производное владение парасфеноидным участком зуба (рис. 2d, f). Aenigmachannidae отличаются от всех других Labyrinthici следующими аутапоморфиями: (а) очень длинная верхнечелюстная кость, выходящая каудально за передний край преоперкли (рис.1b, c, 3a), (b) наличие заметного заглазничного отростка на верхней челюсти (рис. 1c, 3a), (c) фронтальный шов с парасфеноидом, образующим полную межглазничную перегородку (рис. 2f), (d) уникальное количество 29–32 брюшных и 29–31 хвостовых позвонков (рис. 1b), (e) серия из пяти срединных передорсальных костей (надраневральных или безлучевых птеригиофоров) перед спинным плавником (f) 83–85 чешуек в боковая серия; ж) большое количество 40–44 лучей анального плавника (рис. 1а, б). Еще он отличается от всех Anabantoidei и Channidae коротким плавательным пузырем (рис.1д), не доходя до паргипуральной области и отсутствием наджаберной полости и органа (рис. 2h, j). У Aenigmachannidae есть длинные носовые трубки (рис. 1а), циклоидная чешуя, отсутствие шипов в спинном и анальном плавниках (рис. 1а), увеличение числа позвонков, единственное расширение заднего плавательного пузыря в сочетании с абдоминализацией передних десяти постанальных позвонков (рис.1) и пяти жаберных лучей, но отличаются от них аутосфенозом, который исключен из крыши черепа, а лобная часть широко сшита только с птеротиком (рис.2a – f), простейшие и базиокципиталы, образующие равные части буллы для саккулита (рис. 2b, c), наличие множества хвостовых позвонков и, следовательно, отсутствие абдоминализации заднего позвоночного столба (рис. 1b), и отсутствием крючковидного отростка метаптеригоида (рис. 3а), отсутствием кости Дей (рис. 1г) и канала боковой линии тела.
Рисунок 3Очищенные и окрашенные челюсти и нижнебедная дуга сбоку ( a ) Aenigmachanna gollum , (KUFOS 2019.8.226) 60,6 мм, ( b ) Parachanna africana , (MTD-F39824) 150 мм и ( c ) Channa punctata , (MTD-F39825) 118 мм. Обратите внимание на наличие верхнечелюстных отростков (отмечены красными стрелками) в ( a ) и их отсутствие в ( b ) и ( c ), наличие клыковидных зубов (отмеченных черными стрелками) в зубной части ( b ). ) и ( c ) и их отсутствие в ( a ) и наличие длинного крючковидного отростка (отмечено черными стрелками) на метаптеригоиде в ( b ) и ( c ) и его отсутствие в ( a ). ).
Aenigmachannidae: морфологически плезиоморфная линия змееголов
Удлиненное тело, длинные спинной и анальный плавники и передние ноздри без шипов, заметно расширенные в виде носовых трубок (рис. 1a). в этих внешних особенностях. Однако детальное изучение его внутренних морфологических признаков выявляет большое количество существенных отличий. Как показал наш морфологический анализ, многие из них представляют собой более примитивные условия, чем те, которые встречаются у любого из видов чаннов (рис.5, дополнительные фиг. S1, S2) и обсуждается ниже. Монофилия семейства Channidae ранее была основана на нескольких предполагаемых синапоморфиях 31,32 , для которых мы исследовали состояние персонажа в Aenigmachanna (число в скобках после символа относится к номеру символа в дополнительном материале).
а. Отический булл для саккулита в основном содержится в протозе (21)
У Aenigmachanna прототическая часть буллы не увеличена в размерах (рис.2г, е) и вмещает лишь около половины саккулита (рис. 2г). Это состояние встречается во всех других членах Labyrinthici, не являющихся каналом, и представляет собой примитивное состояние символа.
г. Метаптеригоид с антеродорсальным крючковатым отростком, приближающимся к нейрокранию или сочленяющимся с ним (33)
Aenigmachanna лишен крючковидного отростка метаптеригоида ченноглазых (Рис. 3). Такой процесс также отсутствует во всех не-канальных лабиринтах и, как правило, в костистых костях и, таким образом, представляет собой еще одно примитивное состояние в Aenigmachanna .
г. Дополнительные органы дыхания с респираторными узелками на первом и втором наджаберных, поджаберных и парасфеноидных (30)
Aenigmachanna полностью лишены наджаберной полости и наджаберного органа (рис. 2g – j), а также отсутствуют респираторные узелки на наджаберных и надчелюстных суставах. или парасфеноид, опять же более примитивное состояние характера, чем у Channidae. Отсутствие наджаберной полости и лабиринтного органа у Aenigmachanna может указывать на то, что наджаберные полости и органы чванов и анабантоидов развились независимо или что Aenigmachanna потеряли их вторично.Детальное исследование жаберных кровеносных сосудов в Aenigmachanna , которые разделяют синапоморфные условия у каналов и анабантоидов, может предоставить доказательства для выбора между этими двумя разными гипотезами.
г. Присутствие ‘Day’s bone’ sensu
31 , аутогенной удлиненной косточки (как отделившаяся гемальная шипа) между гемальными шипами PU2 и PU3 (15)Дневная кость отсутствует у Aenigmachanna (рис. 1d), что однако имеет раздвоенный гемальный шип в предуральном центре 2.С точки зрения своего положения передняя часть этой разветвления у Aenigmachanna находится в положении кости Дей у Channidae и может представлять более примитивное состояние, при котором она еще не является аутогенной, если только она не слита онтогенетически в Aenigmachanna . к гемальному отделу позвоночника. Это может означать, что кость Дэя в каналах потенциально образовалась из отслоившейся передней части раздвоенного гемального шипа на предуральном центре 2.
e. Отсутствие шипов на всех плавниках (3,5,8)
Aenigmachanna разделяет с Channidae производное отсутствие шипов спинного и анального плавников, которое сочетается у обоих таксонов с очень длинными спинными плавниками с мягкими лучами (> 30 лучей) и анального (> 20 лучей) плавников (рис.1а). Тазовые плавники, которые также бесхребетны у чанвидов, отсутствуют у Aenigmachanna .
ф. Наличие внутреннего ряда клыковидных зубов и внешнего ряда очень маленьких зубов по длине кости (37)
Мюррей 32 добавило этот признак как дополнительную предполагаемую синапоморфию каннелитов (проиллюстрировано здесь на рис. 3b, c). Aenigmachanna не имеет более крупных клыковидных зубов каналов в зубной части (сравните рис. 3a с b и c), но имеет ряд конических зубов аналогичного размера (рис.1c, 3a), как и другие лабиринты, не связанные с каналом, опять же более примитивное состояние.
Мы заключаем, что Aenigmachanna не разделяет пять из шести синапоморфий канала 31,32 , а скорее имеет более примитивное состояние характера по сравнению с таковым у Channidae.
Для дальнейшей оценки филогенетического положения Aenigmachanna по отношению к Channidae и другим Labyrinthici мы провели экономичный анализ 46 морфологических признаков и 15 таксонов Labyrinthici, результаты которого показаны на рис.5. Aenigmachanna восстанавливается как сестринская группа семейства Channidae, положение, выделенное рядом дополнительных примитивных символов, которое Aenigmachanna разделяет с неканнидными Labyrinthici, как показано ниже.
г. Наличие многочисленных хвостовых позвонков (11)
Виды родов Parachanna и Channa очень необычны среди костистых позвонков тем, что имеют удлиненные тела с 40–66 позвонками, из которых, однако, только самые задние 4–6 морфологически являются наиболее необычными. хвостовые позвонки с гемальными шипами (рис.4). Это сочетается с наличием парапофизов и ребер на всех позвонках впереди этих 4-6 истинных хвостовых позвонков. Такая абдоминализация большей части постанального позвоночника является синапоморфией Channidae. Члены их сестринской группы, Anabantoidei, как и все остальные Labyrinthici, лишены абдоминализации задней каудальной области и имеют типичное отделение брюшной полости от хвостового позвоночного столба примерно посередине тела (Рис. 4). Aenigmachanna (рис.1b, 4), а также у всех неканальных Labyrinthici есть примитивное состояние позвоночного столба, разделенного на брюшной и каудальный отделы примерно одинаковой длины (29–32 брюшных и 29–31 хвостовых позвонков у Aenigmachanna ). Задние 10–12 брюшных позвонков в Aenigmachanna , однако, расположены кзади от ануса и начала анального плавника (рис. 1, 4), но имеют парапофизы и ребра и не имеют гемальных шипов и, следовательно, представляют собой позвонки передней части брюшной полости. постанального позвоночника.Позвонки, расположенные в том же положении, что и начало ануса / анального плавника, у некванвидных лабиринтов и у генерализованных перкоморфов являются хвостовыми позвонками (рис. 4).
Рисунок 4Вид сбоку скелета обобщенного перкоморфа Поздний ( a ), анабантоид Ctenopoma ( b ), энигмаханнид Aenigmachanna ( c ) и канал Parachanna Parachanna ( д ). Зеленая стрелка отмечает положение вентиляционного отверстия, красной стрелкой — положение первого хвостового позвонка (самый передний позвонок с гемальным остью).Обратите внимание на более или менее равное разделение позвоночного столба у , поздний ( a ) и Ctenopoma ( b ) на брюшной и хвостовой позвонки ( a , b , d адаптировано из 33 ) с расположением ствола и первого хвостового позвонка близко друг к другу, а хвостовой и постанальный отдел позвоночника одинаковой длины. Обратите внимание на частичную абдоминализацию постанального позвоночника в Aenigmachanna и почти полную абдоминализацию в Parachanna .
ч. Одиночный плавательный пузырь, не доходящий сзади до паргипурального пузыря, но оканчивающийся в середине тела (17)
С абдоминализацией каудального позвоночного столба у Channoidei связан расширенный назад плавательный пузырь, который достигает паргипурального уха в Channa и Параханна 34,35 . Расширение плавательного пузыря аналогичной длины также присутствует у Anabantoidei, но здесь удлинение состоит из парных расширений, идущих кзади вдоль левой и правой стороны гемальных шипов хвостовых позвонков 34 .Расширение заднего плавательного пузыря до паргипурального отдела было предположено как чанно-анабантоидная синапоморфия 31 . Наличие более короткого плавательного пузыря у Aenigmachanna (рис. 1e), который заканчивается на уровне 8-го постанального позвонка, отличает этот таксон от всех других каналов и анабантоидов. Это состояние характера интерпретируется как примитивное состояние, присущее nandid, badid и pristolepidid Labyrinthici и другим перкоморфам, при этом удлинение заднего плавательного пузыря развивалось независимо у анабантоидов и канноидов.Такая интерпретация может также объяснить различие в характере удлинения (одиночное удлинение средней линии у каналов, парные дивертикулы у анабантоидов).
и. Аутосфенотический канал не является частью крыши черепа и не несет канал боковой линии (19,20)
Каналы необычны среди костистых кости тем, что височный канал, обычно ограниченный лобными и птеротическими костями дермы, проходит в аутосфенозе, хондральном отделе. кость, которая также является частью крыши черепа (рис. 2а). У Aenigmachanna и других лабиринтозов, не являющихся каналом, аутосфенотик исключен из крыши черепа, а височный канал по боковой линии проходит исключительно в птерозе и лобной части, которые находятся в прямом контакте (рис.2б).
Дж. Базиокципитал не имеет парных фланцев или отростков (22)
Каналы имеют пару удлиненных боковых фланцев или круглых мыщелков, которые образуют сочленение с верхней глоточной челюстью через суставную поверхность на глоточно-жаберных челюстях 2 + 3. мыщелки и верхняя глоточная челюсть не сочленяются с основной затылочной. Это потенциально еще одно плезиоморфное состояние в Aenigmachanna . Отсутствие таких отростков также может быть интерпретировано как вторичная потеря у Aenigmachanna , потому что анабантоиды имеют парные базиоциципитальные глоточные отростки, которые могут быть гомологичны парным отросткам у каналов.Все вышеупомянутые гипотетические примитивные состояния признаков в Aenigmachanna и их апоморфное состояние у Channidae предполагают, что они являются сестринскими группами и что Aenigmachannidae представляют собой линию, которая отделилась от других змееголовых рыб до того, как они разделились на африканский род Parachanna и азиатский род. Чанна . Среди Labyrinthici, Channidae и Aenigmachannidae имеют общие производные состояния следующих признаков, как предполагаемые чановые синапоморфии (см. Символы 1–5, 7, 9, 10, 16, 41 в дополнительном файле): длинные носовые трубки, циклоидные чешуйки, отсутствие дорсальных — и шипы анального плавника, большое количество лучей спинного и анального плавников, увеличение числа позвонков и одно расширение заднего плавательного пузыря в сочетании с абдоминализацией, по крайней мере, переднего постанального позвоночника и только пятью жаберно-стегальными лучами.
Aenigmachannidae как отдельная линия змееголовых: оценки времени молекулярной дивергенции и биогеографические последствия
Наш анализ двух различных наборов молекулярных данных, один из которых основан на полных митохондриальных геномах, поддерживает точку зрения, что Aenigmachanna gollum образует линию, отделены от Channa и Parachanna на долгое время. В зависимости от проанализированного набора данных (дополнительные рисунки S3, S4) Aenigmachanna определяется либо как сестринская группа для Channidae (набор данных 2), либо как сестринская группа для Channa (набор данных 1).Эти альтернативные размещения в дополнение к альтернативному положению Aenigmachanna в качестве сестринской группы по отношению к Parachanna , однако, не могли быть отклонены статистически (дополнительный рис. S3), что указывает на то, что трихотомия в корне дерева не может быть решена в дальнейшем. с этими наборами данных. Некоторые короткие междоузлия, расположенные глубоко в филогенезе, как известно, трудно разрешить даже при применении наборов данных в масштабе генома 36,37 . Здесь спорными филогенетическими отношениями может быть даже горстка генов, что затрудняет наши поиски разрешения бесчисленных ветвей древа жизни 38 .Хотя один из наших наборов данных был основан на полной митохондриальной ДНК, только больше данных в филогеномном масштабе может помочь лучше разрешить топологическую неопределенность относительно филогенетического положения Aenigmachanna . Несмотря на такой уровень неопределенности, результаты обоих анализов подчеркивают уникальность линии передачи Aenigmachanna и ее долгое отделение от линий передачи Parachanna и Channa .
Однако наш морфологический анализ однозначно относит Aenigmachanna к сестринской группе по отношению к Channidae (рис.5), что также подчеркивается большим количеством морфологически примитивных признаков Aenigmachanna по отношению к Channidae (рис. 5). Учитывая, что это размещение Aenigmachanna дополнительно подтверждается результатами анализа набора данных 2 и не отклоняется набором данных 1, мы использовали ограниченное дерево на основе набора данных 1, отражающего родственные связи двух таксонов, для выполнения оценка времени расхождения (рис. 6). Мы выбрали набор данных 1 вместо набора данных 2, поскольку первый позволил нам выполнить калибровку на основе ствола и короны двух ключевых ископаемых, которые мы использовали.Мы подсчитали, что Aenigmachanna отделились от сестринской группы, Channidae, по крайней мере, примерно на 109 млн лет назад (диапазон 83–136 млн лет назад) при использовании калибровки коронки и 34 млн лет назад (диапазон 33–35 млн лет назад) при калибровке стержня (рис. 6).
Рис. 5Филогенетические отношения Aenigmachannidae (отмечены красным) на основе 46 морфологических признаков. Показано дерево строгого консенсуса двух самых экономных деревьев (длина дерева = 64 шага, индекс согласованности (CI) = 0,72, индекс удержания (RI) = 0.89). Числа в узлах соответствуют значениям начальной загрузки (1000 повторов). Плезиоморфные состояния характера Aenigmachanna по отношению к Channidae, обсуждаемым в тексте, показаны на дереве с соответствующими номерами.
Рис. 6Оценки времени дивергенции для Aenigmachannidae (отмечены красным) и других Labyrinthici на основе молекулярных данных с использованием ограниченной топологии, соответствующей морфологической топологии. Показаны калибровка группы короны (слева) и группы ствола (справа) окаменелостей (C1: † Osphronemus goramy ; C2: † Parachanna fayumensis ).
Уникальная ветвь Aenigmachannidae может также занимать ключевую позицию среди Channoidei для нашего понимания биогеографии змееголова и эволюционного разнообразия, учитывая его предполагаемый возраст. Применяя калибровку короны, диапазон нашей минимальной оценки возраста 83–136 миллионов лет для линии Aenigmachanna возвращает его к периоду, когда суперконтинент Гондвана начал распадаться, а Индия / Мадагаскар отделились от Африки примерно на 120 миллионов лет. тому назад.Это указывает на то, что присутствие Aenigmachanna и Channa в Азии и Parachanna в Африке является результатом наместничества, как недавно было выдвинуто предположение для Channidae 30 . С альтернативной калибровкой ствола и результирующим минимальным возрастом 33–35 миллионов лет сценарий викариации Гондваны кажется менее вероятным, хотя все еще возможен. Альтернативу расселения из Африки в Индию или наоборот после закрытия Тетиса можно исключить, так как самая старая окаменелость канала, африканская † Parachanna fayumensis , предшествовала этому закрытию и образованию сухопутных мостов между азиатским и африканским континентами. .Третья альтернатива для объяснения присутствия представителей Channoidei как в Африке, так и в Азии — это трансокеанское расселение на большие расстояния. Однако есть веские биологические аргументы, противоречащие такому сценарию. В дополнение к тому, что чанны являются основными пресноводными рыбами, виды рода Parachanna , а также все базальные линии рода Channa имеют хорошо развитую родительскую заботу, в которой плавучие икры охраняются у поверхности воды. ; производное ротовой полости развилось только в группе Гачуа в пределах Channa .Из соображений экономии следует выдвинуть гипотезу о том, что эта защита плавающих яиц представляет собой примитивные условия размножения для каннелитов в целом. Если согласиться с этим, то расселение, требующее многочисленных поколений воспроизводящихся взрослых особей, вынужденных жить в морских водах с таким репродуктивным поведением, кажется невозможным, и родительская забота с использованием плавающих яиц также неизвестна ни одному из видов морских костистых. Это оставляет в качестве наиболее правдоподобного сценария для объяснения чанноидного афро-азиатского распространения гондванское наместничество, в котором линии передачи Aenigmachanna и Channa присутствовали на дрейфующей на север Индийской плите.
Существование такой древней уникальной линии, сохранившей большое количество плезиоморфных условий по отношению к более богатой видами сестринской группы, не является чем-то необычным среди рыб. Stiassny & de Pinna 2 представили примеры из пресноводных рыб, которые они назвали «базальными таксонами», то есть филогенетически и морфологически примитивными таксонами, которые имеют уникальное значение для понимания эволюции сестринской группы, зачастую богатой видами. Они также заметили, что эти «базальные таксоны» часто имеют ограниченное распространение, что делает их очень восприимчивыми к изменениям среды обитания и экологическому давлению.Это также относится к Aenigmachanna , подземная среда обитания которого находится под сильным влиянием изменений уровня грунтовых вод в густонаселенных частях прибрежных латеритных районов, где в водоносные горизонты штата выходит более 6 миллионов приусадебных колодцев. 39 .
Открытие новых видов рыб, таких как Aenigmachanna gollum и A. mahabali , которые нельзя однозначно отнести к известным таксонам на уровне семейства, является редким, но за последние восемь лет появилось три новых семейства костистых, все монотипные. , были описаны для приспособления к морфологически весьма необычным костистым рыбам: ангуиллиформной Protanguilla palau 3 , сомику Kryptoglanis shajii 14 и характерной Tarumania walkerae 40 .Наряду с этими тремя таксонами Aenigmachanna , по нашему мнению, является одним из самых интересных ихтиологических открытий последнего десятилетия, и необходимы дальнейшие подробные исследования, чтобы полностью понять его анатомические характеристики и основные особенности его биологии, особенно его воспроизводства. а также его потребности в среде обитания и распространение в Южной Индии.
ВАДХАВАН Д., ВАЛИЯ ВК: Три новых вида из
П О Л И С Й О У Р Н А Л О Ф Е Н Т О М О Л О Г И П О Л С К И Е П И С М О Е Н Т О М О Л О Г И С З Н Е ТОМ.76: 207-219 Быдгощ, 30 сентября 2007 г. Три новых вида рода Torodora Meyrick (Lepidoptera: Lecithoceridae: Torodorinae ) из Северо-Западной Индии ДИПАК ВАДХАВАН *, ВИРИНДЕР КУМАР ВАЛИЯ ** * Отдел энтомологии, факультет зоологии, Пенджабский университет, Чандигарх — 160014 , ИНДИЯ ** Отдел энтомологии, отделение зоологии, Пенджабский университет, Чандигарх — 160014, ИНДИЯ, электронная почта: virinder_k_walia @ rediffmail.com АННОТАЦИЯ. Три новых северо-западных вида из рода лецитоцеридов Torodora MEYRICK подсемейства Описаны Torodorinae. КЛЮЧОВІ СЛОВА: Lecithoceridae, Torodorinae, Северо-Западная Индия, Torodora, три новых вида . ВВЕДЕНИЕ Род Torodora был создан на основе типа — вида , характеризующего MEYRICK из Бирмы под семейством Gelechiadae (MEYRICK 1894).Это семейное название первоначально записывалось как Gelechidae (STANTON 1854), впоследствии было исправлено как Gelechiidae (NYE & FLETCHER 1991). CLARKE (1955) относился к Torodora как к семейству Timyridae, позже названному NYE & FLETCHER (1991) синонимом под Lecithoceridae. До этого GOZMANY (1978) предложил подсемейство Torodorinae в составе семейства Lecithoceridae с родом Torodora в качестве типа. Этот род представлен 85 видами со всего мира (PARK & HEPPNER 2000).Из них 82 известны из Восточного региона, 2 — из Палеарктики и только 1 — из Эфиопского региона (PARK & HEPPNER 2000). На данный момент 12 видов были зарегистрированы под Torodora из Индии. (MEYRICK 1911, GAEDE 1937, ROSE & PATHANIA 2003).
Барлоу, Х.С. (1982) Знакомство с мотыльками Юго-Восточной Азии. Куала-Лумпур, автор. Браун, Ф.Г. (1968) Вредители и болезни леса Плантационные деревья: аннотированный список основных видов, встречающихся в Британское Содружество. Оксфорд: Clarendon Press. Картер, Д.Дж. (1984) Вредитель Lepidoptera Европы с особым упором на Британские острова. Серия Entomologica 31. Гаага: W Junk. Картер, Д.Дж. И Харгривз Б. (1986) Поле Путеводитель по гусеницам бабочек и мотыльков в Великобритании и Европе. Лондон: Коллинз. Chao, C.-L. (1981) Вертикальное распределение лимфатические мотыльки на горе Эмей, включая описание нового вида. Sinozoologia 18: 97-104. Chao, C.-L. (1983) Описание двух новых видов род Numenes из Китая (Lepidoptera: Lymantriidae). Акт. Синица 26 : 452-456. Chao, C.-L. (1985) О четырех новых видах рода Pida. из Китая (Lepidoptera: Lymantriidae). Акт. Синица 28 : 99-103. Chew, K.L. И Киртон, Л. (в печати) Мотылек гусеница, вызывающая высыпания. Малазийский натуралист. Чей, В.К. (1987) Дефолиаторы насекомых леса плантации деревьев в Сабахе. Публикация Центра исследований леса Сабаха 32. Chey, V.K. (1994) Сравнение биоразнообразия между тропическим лесом и плантациями в Сабахе с использованием мотыльков в качестве индикаторов. Неопубликованные Д.Фил. диссертация, Оксфордский университет. Чей, В.К. (1996) Лесные насекомые-вредители в Сабахе. Сабах Forest Record 15. Chey, V.K., Holloway, J.D. & Speight, MR. (1997) Разнообразие бабочек в лесных плантациях и естественных лесах Сабаха. Бык. энт. Res. 87 : 371-385. Collenette, C.L. (1932) Lymantriidae малайских Полуостров. Новит зоол. 38 : 49-102. Колленетт, К.L. (1947) Лимантрииды целебес. Ann. Mag. физ. Hist. (11) 14 : 1-60. Collenette, C.L. (1949a) Лимантрииды Явы. Ann. Mag. физ. список. (12) 1 : 685-744. Collenette, C.L. (1949b) Лимантрииды Бали. Энтомолог. 82 : 169-175. Collenette, C.L. (1953) Новое и малоизвестное Индонезийские Lymantriidae (Lep.). Treubia, 22 : 225-232. Common, I.F.B. (1990) Бабочки Австралии. Мельбурн University Press. Далл Аста, США (1981a) Revision des espces du genre Rahona Griveaud de l’Afrique Centrale (Чешуекрылые: Lymantriidae). Rev. Zool. AFR . 95 : 253-282. Далл Аста, У. (1981b) Nouveaux genres et nouvelles espces de Lymantriidae africains (Lepidoptera). Rev. Zool. афр. 95 : 733-764. Далл Аста, У. (1982) Les espces du genre Lemuriana Griveaud de l’Afrique Centrale (Lymantriidae, lepidoptera). Rev. Zool. афр. 96 : 11-30. Далл Аста У. (1988) Барабанные органы Лимантрииды (Lepidoptera). Nota lepid. 11 : 169-176. Далл Аста У. (1983) Revision du genre Eudasychira Mschler en Afrique Centrale (Чешуекрылые: Lymantriidae). Rev. Zool. афр. 97 : 11-44. Eecke, R. van (1928) De Heterocera van Sumatra — V. Zool. Медед. Лейден 11 : 49-145. Фергюсон, округ Колумбия (1978) Noctuoidea, Lymantriidae. Бабочки из Америки к северу от Мексики. Т. 22-2. Лондон: Э. В. Класси. Фишер, Р.А., Корбет, А.С. И Уильямс, К. (1943) Связь между количеством видов и количеством особей. в случайной выборке из популяции животных. .J. аним. Ecol. 12 : 42-58 . Gardner, J.C.M. (1938) Незрелые стадии индийского Чешуекрылые (1). Lymantriidae. Индийские лесные записи (Н.С.) 3 : 187-212. Хеппнер, Дж. Б. (1991) Фаунистические регионы и разнообразие чешуекрылых. Тропический Lepidoptera 2 (Дополнение 1): 1-85. Heppner, J.B. & Inoue, H. (1992) Lepidoptera of Taiwan , Volume 1, часть 2: Контрольный список, Гейнсвилл, Флорида: Ассоциация тропиков. Чешуекрылые и научные издательства. Холлоуэй, Дж. Д. (1976) Бабочки Борнео с особым упором на Mt.
Kinabalu . Куала-Лумпур: Малайское общество природы. Холлоуэй, Дж. Д. (1982a) Общее размещение Phalaena (Bombyx) pudibunda L. и Phalaena (Bombyx) fascelina L.(Lep: Lymantriidae). Поступления и Труды Британского общества энтомологии и естествознания , 15: 44. Холлоуэй, Дж. Д. (1982b) Таксономическое приложение. В: Barlow, H.S. Введение
к мотылькам Юго-Восточной Азии, с. 174-271. Куала-Лумпур: автор. Холлоуэй, Дж.Д. (1984) Более крупные бабочки национального парка Гунунг Мулу; а
предварительная оценка их распространения, экологии и потенциала как
экологические показатели. В: Jermy, A.C. & Kavanagh, K.P. (ред.) Gunung
Национальный парк Мулу, Саравак, часть II, , с. 149–190. Журнал музея Саравака 30, Special Issue 2. Холлоуэй, Дж. Д. (1989a) Мотыльки. В: Lieth, H & Werger, M.J.A (ред.) Tropical
Экосистемы тропических лесов мира , 14B: 437-453. Амстердам: Эльзевир. Холлоуэй, Дж. Д., Брэдли, Дж. Д. и Картер, Д. Дж. (1987) Lepidoptera. Руководства CIE по насекомым, важным для человека, Том 1 (под ред. К.Р. Беттса). Уоллингфорд: CAB International. Хорсфилд, Т. и Мур, Ф. (1859 [1860]) Каталог Чешуекрылые насекомые в Музее естественной истории Восточно-Индийского дома. Т. II. Лондон: W.H. Аллен. Hutacherern, C. & Tubtim, N. (1995) Контрольный список лесных насекомых в Тайланде. Бангкок: Управление экологической политики и планирования. Иноуэ, Х., Суги, С., Куроко, Х., Мориути, С., Кавабе, А. (1982) Мотыльки Японии. 2 тт. Токио: Коданша. Kalshoven, L.G.E. (1981) Вредители сельскохозяйственных культур в Индонезии. Пересмотренный и переведен П.А. ван дер Лаан с Г.Х.Л. Ротшильд. Джакарта: P.T. Ихтиар Бару-ван Хов. Кишида Ю. (1993) Lymantriidae in Moths of Nepal 2. Tinea 13 (Прил.3): 80-95. Kishida, Y. (1994) Lymantriidae ( Euproctis with S. Sugi) у бабочек Непала 3. Tinea 14 (Дополнение 1): 72-82 Kishida, Y. (1995) Lymantriidae in Moths of Nepal 4. Tinea 14 (Приложение 2): 44-46. Кишида Ю. (1996) Neocifuna Inoue, 1982, младший синоним Ilema Мур, [1860] (Lymantriidae). Japan Heterocerists J. 188: 212-213. Китчинг, И.Дж. И Роулинз, Дж. Э. (1999 [1998]) The Noctuoidea. В: Кристенсен, Н.П. (ред.) Чешуекрылые, мотыльки и бабочки Том 1: Эволюция, систематика и биогеография. С. 355-401 . Справочник по зоологии IV (35). Берлин и Нью-Йорк: Вальтер де Грюйтер. Kobes, L.W.R. (1985) Thyatiridae, Agaristidae и Noctuidae (часть 1: Pantheinae и Catocalinae) Суматры. Heterocera Sumatrana 4. Куроко, Х. и Левванич, А. (1993) Чешуекрылые тропические вредители Фруктовые деревья в Таиланде. Токио: Агентство международной корпорации Японии. Lemaire, C. & Minet, J. (1999 [1998]) Bombycoidea и их родные. В: Кристенсен, Н.П. (ред.) Чешуекрылые, мотыльки и бабочки Том 1: Эволюция, систематика и биогеография. С. 321-353. Справочник по зоологии IV (35). Берлин и Нью-Йорк: Вальтер де Грюйтер. Мабберли, Д.Дж. (1987) Книга растений, портативный словарь высшие растения. Издательство Кембриджского университета. Mackey, A.P. (1981) Обзор новогвинейских родов Parakanchia Bethune-Baker и Soretoneura Collenette (Lepidoptera: Lymantriidae). .J. Aust. энт. Soc. 20: 119-126. Mackey, A.P. (1984) Примечания к некоторым типам Toxopeus Redoa (Lepidoptera: Lymantriidae). Восточные насекомые 18 : 5-12. Maes, K. (1984a) Сравнительное исследование гениталий некоторых Lymantriidae Hampson, [1893] из Палеарктики. Mededel. К. акад. Wetenschappen Belg . 46 : 125-149. Maes, K. (1984b) Некоторые замечания по роду Euproctis Hbner, [1819] 1816 г. (Lepidoptera: Lymantriidae). Nota lepid. 7 : 55-58. Mann, D.G. (1998) Эренбергиана: проблемы неуловимых типов и старые коллекции с особым упором на диатомовые водоросли. Линней, Особый Выпуск 1: 63-88. Мэтьюз М. (1991) Классификация Heliothinae. Натуральный Бюллетень института ресурсов 44. Миллер, Дж. (1991) Кладистика и классификация Notodontidae (Lepidoptera: Noctuoidea) по морфологии личинок и взрослых особей. Бык. Амер. Mus. физ. Hist., 204. Minet, J. (1998) Некоторые актуальные вопросы филогенетики Macrolepidoptera систематика [аннотация представленной статьи]. Программа SEL XI съезда Лепидоптерология, , с. 72-73. Mitchell, A. (1998) Систематика совок и родственных бабочек (Lepidoptera: Noctuidae) выведены из генов, кодирующих ядерные белки. к. Д. докторскую диссертацию в Мэрилендском университете. Мур, Ф. (1883) Чешуекрылые Цейлона. Том 2. Лондон: Рив и Ко. Мерфи, Д.Х. (1990) Естественная история насекомых. травоядные растения на мангровых деревьях в Сингапуре и его окрестностях. Raffles Bull. Zool. 38 : 119-203. Нильсен, Э.С., Эдвардс, Э.Д. & Rangsi, TV. (ред.) (1996) Контрольный список чешуекрылых Австралии. CSIRO, Австралия. Pholboon, P. (1965) Список хозяев насекомых Таиланд. Бангкок: Министерство сельского хозяйства Таиланда и операции в США Миссия в Тайланд. Пул Р.В. (1995) Noctuoidea, Noctuidae (частично). В: Доминик, Р. Б. и др. Бабочки Америки к северу от Мексики, 26.1. Вашингтон: Фонд энтомологических исследований клина. Riotte, J.C.E. (1979) Австралийская и папуасская кочка бабочки комплекса Orgyia (Lepidoptera: Lymantriidae). Тихоокеанские насекомые 20 : 293-311. Робинсон, Г.С. (1975) Макролепидоптеры Фиджи и Ротума, таксономическое и географическое исследование. Фарингдон: Клесси. Schintlmeister, A. (1994) Аннотированный и иллюстрированный контрольный список Lymantriidae Суматры с описанием новых виды (Lepidoptera, Lymantriidae), Heterocera Sumatrana 7 : 113-180. Скобл, М.Дж. (1992) Чешуекрылые, форма, Функция и разнообразие. Оксфорд: Издательство Оксфордского университета. Севастопуло, Д.Г. (1938) Ранние этапы индийского Чешуекрылые. Часть 1. J. Бомбей. физ. Hist. Soc. 40: 817-827. Севастопуло, Д.Г. (1940) О пищевых растениях Индии. Bombyces (Heterocera). J. Bombay. физ. Hist. Soc. 41: 817-827. Севастопуло, Д.Г. (1942) Ранние этапы индийского Чешуекрылые.Часть 10. J. Bombay. физ. Hist. Soc. 43: 409-415. Севастопуло, Д.Г. (1943) Ранние этапы индийского Чешуекрылые. Часть 11. J. Bombay. физ. Hist. Soc. 44: 78-87. Севастопуло, Д.Г. (1946) Ранние этапы индийского Чешуекрылые. Часть 15. J. Бомбей. физ. Hist. Соц . 46: 253-269. Speidel, W., Fnger, H. & Naumann C.M. (1996) филогенез совок. Syst.Ent. 21: 219-251. Sugi, S. (ed.) (1987) Личинки больших бабочек в Япония. Токио: Коданша. Toxopeus, L.J. (1948) Заметки о Lymantriidae с частичная ревизия рода Redoa Wlk. (Lep. Heterocera). (Результаты Третья экспедиция Арчболда (1938-1939). Treubia 19: 429-481. Твиди, M.W.F. И Эммет, А. (1991) Отдыхая поза чешуекрылых. В: Эммет, А.М. и Хит, Дж. (Ред.) Мотыльки и Бабочки Великобритании и Ирландии, 7 (2): 46-60. Колчестер: Харли Букс. Ван. H.Y. (1993) иллюстрации Lymantriidae на Тайване. Watson, A. Fletcher, D.S. & Nye, I.W.B. (1980) Родовые названия бабочек мира. Том 2. Лондон: Британский музей. (Естественная история). Worms, C.G.M. de (1979) Lymantriidae. В: Хит, Дж.И Эммет, А. (eds) Мотыльки и бабочки Великобритании и Ирландия, 9 : 66-78. Лондон: Карвен. Юнус, А. и Хо, Т.Х. (1980) Список экономических вредители, растения-хозяева, паразиты и хищники в Западной Малайзии (1920–1978). Министерство of Agriculture, Malaysia, Bulletin 152. Zhang, B-C. (1994) Индекс экономически важных Чешуекрылые. Уоллингфорд: CAB International. |
Пересмотр систематики и эволюции патогенности рода Leptospira через призму геномики
Сборник штаммов и полногеномное секвенирование
В это исследование была включена коллекция экологических изолятов из Азии (Япония и Малайзия), Африки (Алжир и остров Майотта, заморский департамент Франции в Индийском океане), Европы (Франция) и Океании (Новая Каледония). Leptospira Изоляты были извлечены из образцов воды и почвы с разных участков с 2008 по 2017 год. За исключением японских изолятов [17], экологические изоляты, указанные в этом исследовании, ранее не описывались.
ДНК всего 90 изолятов секвенировали с использованием технологии Illumina (таблица S1). У 90 штаммов Leptospira средний размер генома составлял 4 128 000 ± 221 345 п.н. Самый большой геном составлял 4 993 538 п.н., принадлежащих штамму Leptospira putramalaysiae SSW20.Наименьшим геномом оказался геном Leptospira fletcheri , штамм SSW15 T , размером 3733663 п.н. Содержание GC геномов в этом исследовании варьировалось от 37,06 до 47,70. Средняя сборка генома содержала 49 ± 50 контигов (таблица S1).
Филогеномика и идентификация 30 новых
Leptospira видовПолные последовательности генома 90 штаммов, описанных в этом исследовании, сравнивали с ранее опубликованными последовательностями генома уже известных видов Leptospira и штамма GWTS # 1, который был ошибочно отнесен к виду Leptospira alstonii [33, 34 ] (Таблица S1).Отметим, что мы секвенировали геном L . idonii штамм Eri-1 T , поскольку последовательность генома не была доступна для этого вида [13]. Кроме того, геном недавно описанного вида Leptospira macculloughii [15] был исключен из дальнейшего анализа в нашем списке референсных геномов как геном L . macculloughii — результат смешанного культивирования L . meyeri и L . Леветти .
Результаты, полученные при попарном сравнении 124 последовательностей генома, суммированы в виде матрицы в таблице S2. Используя отсечение ANI 95%, которое обычно используется в качестве метрики для определения видов бактерий [35], мы установили существование 64 различных видов Leptospira . Эти виды включают 34 ранее описанных вида, 4 новых вида из Японии (Masuzawa et al., ), 25 недавно выделенных видов ( Leptospira kemamanensis sp. Nov., Leptospira andrefontaineae sp. nov., Leptospira bandrabouensis sp. nov., Leptospira bouyouniensis sp. nov., Leptospira congkakensis sp. nov., Leptospira dzianensis sp. nov., Leptospira dzoumogneensis sp. nov., Leptospira fletcheri sp. nov., Leptospira fluminis sp. nov., Leptospira gomenensis sp. nov., Leptospira ilyithenensis sp. nov., Leptospira jelokensis sp.nov., Leptospira kanakyensis sp. nov., Leptospira langatensis sp. nov., Leptospira montravelensis sp. nov., Leptospira mtsangambouensis sp. nov., Leptospira noumeaensis sp. nov., Leptospira ognonensis sp. nov., Leptospira perdikensis sp. ноя, Лептоспира . putramalaysiae sp. nov., Leptospira sarikeiensis sp. nov., Leptospira selangorensis sp. ноя, Leptospira semungkisensis sp. nov., Leptospira koniamboensis sp. nov., Leptospira bourretii sp. nov.) и один новый вид ( Leptospira tipperaryensis sp. nov.), который ранее был ошибочно отнесен к L . alstonii (штамм GWTS # 1 T ) на основе генов 16S рРНК и secY [33, 34]. Только один репрезентативный штамм каждого из 64 видов Leptospira был оставлен для дальнейшего анализа.Названия и описания происхождения этих новых видов указаны ниже. Учитывая нашу тщательно подобранную базу данных, мы оценили таксономическое назначение последовательностей генома, уже доступных в GenBank, с нашими соответствующими эталонными штаммами (файл S1). При этом мы могли легко обнаружить неправильную классификацию штаммов (например, для некоторых L . santarosai , L . weilii и L . опросить присвоенных штаммов, см. Файл S1), которые подтверждают достоверность использование нашей базы данных и методологии.
Описание 2-х и 4-х подкладов
Филогенетическое положение каждого вида было надежно оценено путем выполнения молекулярной филогении на основе 1371 последовательности ортологичных генов, связанных с матрицей значений ANI для каждого из 64 видов Leptospira (рис. 1). Рисунок подтверждает 64 хорошо очерченных вида Leptospira . Значения ANI согласуются с их филогенетическими отношениями. Значения межвидового ANI варьировались от ~ 69% до ~ 94%.
Рис. 1. Филогенетическое дерево, основанное на последовательностях 1371 гена, признанных ортологичными.
Матрица представляет рассчитанные значения ANIb для всех геномных последовательностей. Ветви окрашены в соответствии с их принадлежностью к четырем основным субкладам: P1 (красный), P2 (фиолетовый), S1 (зеленый) и S2 (синий). Значение начальной загрузки указано для одного узла (что соответствует разделению между L . biflexa штамм Patoc1 и L . bouyouniensis штамм 201601297), поскольку все остальные имеют максимальное значение 100.Цветной круг, согласно легенде, представляет географическое происхождение каждого из новых видов, описанных в этом исследовании. Узел 1 указывает узел, от которого происходят патогенные виды, наиболее часто вызывающие болезни человека.
https://doi.org/10.1371/journal.pntd.0007270.g001
Четыре из новых 30 видов, изолированных от естественной среды в Малайзии, Майотте и Новой Каледонии, а также у мелких млекопитающих в Ирландии, были отнесены к происхождение возбудителей болезней.Десять новых видов, выделенных из Малайзии, Майотты, Японии и Новой Каледонии, были идентифицированы как часть промежуточных продуктов. Двенадцать новых видов, выделенных из Малайзии, Майотты, Японии и Новой Каледонии, были отнесены к сапрофитам. Наконец, четыре новых вида, выделенных из Японии, Алжира и Франции, были помещены в сестру клады той, которая образована сапрофитами вместе с L . idonii . Используя это большое количество новых видов, мы смогли уточнить различные клады и идентифицировать две основные клады и четыре субклада в роде Leptospira .Двумя основными кладами являются: «Сапрофиты», содержащие виды, изолированные в естественной среде и не ответственные за инфекции, и «Патогены», содержащие все виды, ответственные за инфекции у людей и / или животных, а также виды окружающей среды, для которых статус вирулентности не был определен. доказано. Каждая из двух клад подразделяется на две субклада. Мы предлагаем новую номенклатуру, чтобы ограничить предположение о характере вирулентности, которые еще предстоит охарактеризовать (рис. 1): клады P и S и субклада P1 (ранее описываемая как группа патогенов), P2 (ранее описываемая как промежуточная группа), S1 (ранее описываемая как группа сапрофитов) и S2 (описываемый здесь новый субклад, который включает L . идонии ).
Мы также сравнили значения аминокислотной идентичности (AAI) из транслированных последовательностей разных видов. Этот анализ дополняет анализ ANI в том смысле, что аминокислоты эволюционируют медленнее, чем нуклеотиды (вырожденность генетического кода), что позволяет визуализировать более крупные и более предковые группы [36]. Как и ожидалось, мы находим те же клады и субклады, что и со значениями ANI (S1 Рис). Однако сигнал сильнее, и клады лучше определены.Аналогичным образом, процент значений консервативных белков (POCP) также был рассчитан для 64 сравниваемых последовательностей генома (S2, фиг.). POCP определяли с использованием всех белков геномов, чтобы сделать вывод о генетическом и фенотипическом родстве между парой видов [37]. С помощью этого анализа мы можем ясно видеть две клады S и P (еще раз подтверждая генетическое родство между «бывшей» кладой сапрофитов S1 и новой субкладой S2).
Фенотипическая характеристика новых видов и нового субклада S2
30 новых видов Leptospira хорошо растут в жидком EMJH при 30 ° C.Под микроскопией в темном поле штаммы этих новых видов подвижны и демонстрируют характерные крючковидные и спиралевидные концы, обусловленные вращением эндофлагеллы. Все они обладают морфологией, соответствующей роду Leptospira , то есть тонкими, длинными и спиралевидными клетками.
Был проведен более подробный фенотипический анализ штаммов, составляющих новый субклад S2. L . кобаяши , L . ilyithenensis , L . idonii и L . ognonensis были протестированы на фенотип роста в различных условиях. Изоляты также были отобраны для получения по одному представителю от каждого из трех других субкладов (P1: L . interrogans штамм L495; P2: L . licerasiae штамм VAR010 T ; S1: L . biflexa штамм Patoc 1). Оптимальная температура для роста 30 ° C для всех видов Leptospira .Время удвоения в жидком EMJH при 30 ° C составляет от 15 до 23 часов, за исключением L . kobayashii со временем удвоения 8 ч. Виды субклада S2 хорошо растут при 14 ° C, но не при 37 ° C, демонстрируя характеристики роста, обычно наблюдаемые для видов субклада S1 (ранее называемые сапрофитами) и не наблюдаемые для видов из клады P, которые также могут расти при 37 ° C. Точно так же виды субклада S2 могут расти в EMJH, дополненном аналогом пурина 8-азагуанином, который обычно использовался в качестве дифференциального агента для разделения патогенных (P1) и сапрофитных (S1) Leptospira (таблица S4).Для оценки вирулентности видов этого нового субклада два репрезентативных изолята ( L , ognonensis и L , ilyithenensis ) вводили хомякам в высокой дозе (10 8 бактерий). Животные, инфицированные этими новыми видами, не проявляли никаких клинических признаков лептоспироза, и бактерии не выделялись из почек или печени инфицированных животных с использованием гомогенатной культуры. Напротив, заражение вирулентным L . interrogans штамм L495 вызвали смерть инфицированных хомяков. Эти результаты указывают на неспособность этих новых видов установить острую инфекцию или почечную колонизацию в этой животной модели.
Морфология клеток была аналогична таковой у представителей рода Leptospira . Клетки имели форму спирали с длиной от 9 до 14 мкм, диаметром ~ 0,2 мкм и длиной волны от 0,6 до 0,9 мкм (фиг. S3 и таблица S4).
Описание Leptospira dzianensis sp.ноя : dzi.an.en’sis. Н.Л. fem. прил. dzianensis Дзиани, озера на Майотте. Типовой штамм — M12A T , выделенный из пробы воды в Дзиани, Майотта. Регистрационный номер генома — RQHR00000000. Геномное содержание G + C типового штамма составляет 45,5%. Принадлежит к субкладу P1.
Описание Leptospira tipperaryensis sp. ноя : tip.pe.ra.ry.en’sis. Н.Л. fem. прил. tipperaryensis, относящийся к Типперэри, графству в Ирландии. Типовым штаммом является GWTS # 1 T , выделенный из Crocidura russula в Типперэри, Ирландия.Ранее присвоено L . alstonii [33, 34], этот штамм относится к субкладу P1. Регистрационный номер генома — GCA_001729245.1. Геномное содержание G + C типового штамма составляет 42,4%.
Описание Leptospira gomenensis sp. ноя : go.men.en’sis. Н.Л. fem. прил. gomenensis из Каала-Гомен, деревни в Новой Каледонии. Типовым штаммом является KG8-B22 T , выделенный из образца почвы в Каала-Гомен, Северная провинция Новой Каледонии. Регистрационный номер генома — RQFA00000000.Геномное содержание G + C типового штамма составляет 46,1%. Принадлежит к субкладу P1.
Описание Leptospira putramalaysiae sp. ноя : put.ra.ma.lay’si.ae. Н.Л. ген. п. putramalaysiae из Путра Малайзия, университет, в котором находится лаборатория, в которой был выделен штамм. Типовой штамм — SSW20 T , выделенный из пробы воды в Сунгай Конгкак, Малайзия. Регистрационный номер генома — RQEQ00000000. Геномное содержание G + C типового штамма составляет 42,5%. Принадлежит к субкладу P1.
Описание Leptospira andrefontaineae sp. ноя : an.dre.fon.tai’ne.ae. Н.Л. ген. n.andrefontaineae Андре-Фонтен, названный в честь Женевьев Андре-Фонтен, французского ветеринара, внесшего значительный вклад в изучение лептоспироза животных. Типовым штаммом является PZF11-2 T , выделенный из пробы воды в Нумеа, Новая Каледония. Регистрационный номер генома — RQEY00000000. Геномное содержание G + C типового штамма составляет 39,9%. Принадлежит к субкладу P2.
Описание Leptospira dzoumogneensis sp. ноя : dzou.mog.ne. en’sis. Н.Л. fem. прил. dzoumognensis из Дзумогне, деревни на Майотте. Типовой штамм — M11A T , выделенный из пробы воды в Дзумогне, Майотт. Регистрационный номер генома: RQHS00000000. Геномное содержание G + C типового штамма составляет 41,0%. Принадлежит к субкладу P2.
Описание Leptospira koniamboensis sp. ноя : Н.Л. fem. прил. koniamboensis из Кониамбо, горы в Новой Каледонии.Типовой штамм — TK1-4 T , выделенный из пробы воды в Коне, Северная провинция Новой Каледонии. Регистрационный номер генома — RQFY00000000. Геномное содержание G + C типового штамма составляет 39,0%. Принадлежит к субкладу P2.
Описание Leptospira sarikeiensis sp. ноя : sa.ri.kei.en’sis. Н.Л. fem. прил. sarikeienis Сарыкейского района. Типовым штаммом является LIMR175 T , выделенный из пробы воды в Сараваке, Малайзия. Регистрационный номер генома — RQGF00000000.Геномное содержание G + C типового штамма составляет 40,3%. Принадлежит к субкладу P2.
Описание Leptospira johnsonii sp. ноя : john.so’ni.i. Н.Л. ген. п. johnsonii of Johnson, названный в честь Рассела Джонсона, американского микробиолога, который разработал среду EMJH, которая обычно используется для культивирования Leptospira . Типовой штамм — E8 T , выделенный из образца почвы в Ибараки, Япония [17]. Регистрационный номер генома — BFAY00000000. Геномное содержание G + C типового штамма составляет 41.3%. Принадлежит к субкладу P2.
Описание Leptospira fluminis sp. ноя : flu’mi.nis. L. gen. п. fluminis реки. Типовым штаммом является SCS5 T , выделенный из образца почвы в Сунгай Конгкак, Малайзия. Номер доступа к геному: RQEV00000000. Содержание G + C в геноме типового штамма составляет 47,7%. Принадлежит к субкладу P2.
Описание Leptospira fletcheri sp. ноя : flet’che.ri. Н.Л. ген. п. fletcheri of Fletcher, названный в честь Уильяма Флетчера, который сообщил о первом случае лептоспироза в Малайзии в 1927 году.Типовой штамм — SSW15 T , выделенный из пробы воды в Сунгай Конгкак, Малайзия. Регистрационный номер генома — RQET00000000. Геномное содержание G + C типового штамма составляет 47,3%. Принадлежит к субкладу P2.
Описание Leptospira semungkisensis sp. ноя : se.mung.kis.en’sis. Н.Л. fem. прил. semungkisensis реки Семунгкис в районе Хулу Лангат штата Селангор, Малайзия. Типовым штаммом является SSS9 T , выделенный из образца почвы в Сунгай Конгкак, Малайзия.Регистрационный номер генома — RQEP00000000. Геномное содержание G + C типового штамма составляет 42,8%. Принадлежит к субкладу P2.
Описание Leptospira langatensis sp. ноя : lan.gat.en’sis. Н.Л. fem. прил. langatensis района Лангат, Малайзия. Типовой штамм — SSW18 T , выделенный из пробы воды в Сунгай Конгкак, Малайзия. Регистрационный номер генома — RQER00000000. Геномное содержание G + C типового штамма составляет 44,8%. Принадлежит к субкладу P2.
Описание Leptospira selangorensis sp. ноя : se.lan.gor.en’sis. Н.Л. fem. прил. selangorensis штата Селангор, Малайзия. Типовой штамм — SSW17 T , выделенный из пробы воды в Сунгай Конгкак, Малайзия. Регистрационный номер генома — RQES00000000. Геномное содержание G + C типового штамма составляет 40,0%. Принадлежит к субкладу P2.
Описание Leptospira ognonensis sp. ноя : og.non.en’sis. Н.Л. fem. прил. ognonensis из Огнона, реки во Франции.Типовой штамм — 201702476 T , выделенный из пробы воды в регионе Бургундия-Франш-Конте, Франция. Регистрационный номер генома: RQHS00000000. Геномное содержание G + C типового штамма составляет 39,7%. Принадлежит к субкладу S2.
Описание Leptospira ilyithenensis sp. ноя : il.yi.then.en’sis. Н.Л. fem. прил. ilyithenensis из Ильитена, деревни, расположенной в Алжире. Типовой штамм — 201400974 T , выделенный из пробы воды в Ильитене, деревне, расположенной в горах Джурджура, Алжир.Регистрационный номер генома — RQHV00000000. Геномное содержание G + C типового штамма составляет 40,5%. Принадлежит к субкладу S2.
Описание Leptospira kobayashii sp. ноя : ko.ba.ya’shi.i. Н.Л. ген. п. kobayashii из Кобаяши, названный в честь Юдзуру Кобаяши, японского врача и микробиолога, который ввел моноклональные антитела для классификации Leptospira . Типовой штамм — E30 T , выделенный из образца почвы в Гифу, Япония [17].Регистрационный номер генома — BFBA00000000. Геномное содержание G + C типового штамма составляет 40,7%. Принадлежит к субкладу S2.
Описание Leptospira ryugenii sp. ноя : ru.ge’ni.i. Н.Л. ген. п. ryugenii из Ryugen, прозванный в честь Ясутакэ Янагихара, японского микробиолога из Университета Сидзуока, внесшего вклад в хемотаксономию и исследование Leptospira . Типовым штаммом является Yh201 T , выделенный из пробы воды в Сидзуока, Япония [17].Регистрационный номер генома — BFBB00000000. Геномное содержание G + C типового штамма составляет 39,9%. Принадлежит к субкладу S2.
Описание Leptospira bandrabouensis sp. ноя : ban.dra.bou.a.en’sis. Н.Л. fem. прил. bandrabouaensis из Бандрабуа, коммуны Майотта. Типовым штаммом является M10A T , выделенный из пробы воды в Бандрабуа, Майотт. Регистрационный номер генома — RQHT00000000. Геномное содержание G + C типового штамма составляет 37,9%. Принадлежит к субкладу S1.
Описание Leptospira noumeaensis sp. ноя : nou.me.a.en’sis. Н.Л. fem. прил. noumeaensis из Нумеи, столицы Новой Каледонии. Типовым штаммом является PZF14-4 T , выделенный из пробы воды в Нумеа, Южная провинция Новой Каледонии. Регистрационный номер генома — RQFK00000000. Геномное содержание G + C типового штамма составляет 38,3%. Принадлежит к субкладу S1.
Описание Leptospira jelokensis sp. ноя : je.lok.en’sis. Н.L. fem. прил. Jelokensis из Jelok, жилого района в Каджанге, откуда был изолирован штамм. Типовой штамм — L5S1 T , выделенный из образца почвы в Сунгай Джелоке, Малайзия. Регистрационный номер генома — RQGR00000000. Геномное содержание G + C типового штамма составляет 38,9%. Принадлежит к субкладу S1.
Описание Leptospira bourretii sp. ноя : Н.Л. ген. п. bourretii из Бурре, названный в честь Анри Дезире Гастона Бурре (1875–1917), врача и микробиолога, который разработал медицинскую микробиологию в Новой Каледонии.Типовым штаммом является PZF7-6 T , выделенный из образца почвы в Нумеа, Южная провинция Новой Каледонии. Регистрационный номер генома — RQFM00000000. Геномное содержание G + C типового штамма составляет 38,2%. Принадлежит к субкладу S1.
Описание Leptospira kanakyensis sp. ноя : ka.na.ky.en’sis. Н.Л. fem. прил. kanakyensis Канаки, название «Новой Каледонии» для канаков. Типовой штамм — TK5-11 T , выделенный из образца почвы в Коне, Северная провинция Новой Каледонии.Регистрационный номер генома — RQFG000000. Геномное содержание G + C типового штамма составляет 38,5%. Принадлежит к субкладу S1
Описание Leptospira kemamanensis sp. ноя : ke.ma.man.en’sis. Н.Л. fem. прил. kemamanensis Кемаманского района. Типовым штаммом является LIMR131 T , выделенный из пробы воды в Теренггану, Малайзия. Регистрационный номер генома — RQGG000000. Геномное содержание G + C типового штамма составляет 38,9%. Принадлежит к субкладу S1.
Описание Leptospira mtsangambouensis sp.ноя : mtsan.gam.bou.en’sis. Н.Л. fem. прил. mtsangambouensis из Мцангамбуа, деревни на Майотте. Типовым штаммом является M2A T , выделенный из пробы воды в Мцангамбуа, Майотт. Регистрационный номер генома — RQHK00000000. Геномное содержание G + C типового штамма составляет 38,2%. Принадлежит к субкладу S1.
Описание Leptospira bouyouniensis sp. ноя : bou.you.ni.en’sis. Н.Л. fem. прил. bouyouniensis из Буйуни, деревни на Майотте. Типовой штамм — M1A T , выделенный из пробы воды в Буюни, Майотт.Регистрационный номер генома — RQHL00000000. Геномное содержание G + C типового штамма составляет 37,1%. Принадлежит к субкладу S1.
Описание Leptospira ellinghausenii sp. ноя : el.ling.hau.se’ni.i. Н.Л. ген. п. ellinghausenii из Эллингхаузена, названный в честь Германа К. Эллингхаузена, американского микробиолога, который разработал среду EMJH, которая обычно используется для культивирования Leptospira . Типовой штамм — E18 T , выделенный из образца почвы в Фукусиме, Япония [17].Регистрационный номер генома — BFAZ00000000. Геномное содержание G + C типового штамма составляет 37,3%. Принадлежит к субкладу S1.
Описание Leptospira congkakensis sp. ноя : cong.kak.en’sis. Н.Л. fem. прил. congkakensis из Конгкака, рекреационного леса в районе Хулу Лангат штата Селангор, Малайзия. Типовым штаммом является SCS9 T , выделенный из образца почвы в Сунгай Конгкак, Малайзия. Регистрационный номер генома — RQGQ00000000. Геномное содержание G + C типового штамма составляет 38.2%. Принадлежит к субкладу S1.
Описание Leptospira perdikensis sp. ноя : per.dik.en’sis. Н.Л. fem. прил. perdikensis из Пердика, название водопада, где был выделен штамм. Типовой штамм — HP2 T , выделенный из пробы воды в Хулу Пердик, Малайзия. Регистрационный номер генома — RQGA00000000. Геномное содержание G + C типового штамма составляет 38,5%. Принадлежит к субкладу S1.
Описание Leptospira montravelensis sp.ноя : mont.ra’vel.en’sis. Н.Л. fem прил. montravelensis из Монравеля, района Нумеа, где был выделен типовой штамм PZF5-3 T , названный в честь французского мореплавателя Тарди де Монравеля (1811–1864), внесшего вклад в картографию Новой Каледонии. Регистрационный номер генома — RQFN00000000. Геномное содержание G + C типового штамма составляет 37,4%. Принадлежит к субкладу S1.
Геномные характеристики четырех различных субкладов
Кроме того, зная, что существует четыре различных филогенетических субклада видов Leptospira , было интересно проверить открытие пангенома между и внутри каждого из субкладов (Рис. 2).Неудивительно, что в этом анализе субклады отличаются друг от друга. Интересно, что субклад P1 содержит наиболее открытый пангеном. Это говорит о большом разнообразии репертуара генов в этом конкретном субкладе. Тот факт, что P1 имеет более открытый пангеном, позволил нам проверить распределение генов для различных групп (рис. 3). Этот анализ показывает, что, в отличие от видов из P1, все группы имеют примерно одинаковое количество (≈1X) кластеров генов в основном геноме (ортологичный ген во всех геномах) по сравнению с кластерами генов, обнаруженными у одного вида (ген присутствует только в одном геноме). разновидность).Внутри субклада P1 наблюдается сильное обогащение (≈4X) кластерами генов, которые уникальны для одного вида (6252) по сравнению с кластерами генов в основном геноме (1560).
Рис. 2.
Эволюция (A) пангенома и (B) кор-генома для геномов четырех лептоспиральных субкладов. Анализы, выполненные с помощью GET_HOMOLOGUES (с использованием 17 геномов для P1, 21 для P2, 21 для S1 и 5 для S2), подчеркивают, что группа P1 имеет более открытый пангеном.
https: // doi.org / 10.1371 / journal.pntd.0007270.g002
Рис. 3.
Распределение пан-генома по четырем категориям (облако, раковина, мягкое ядро и ядро) для видов из субкладов (A) P1, (B) P2, (C) S1 и (D) S2. Анализы, выполненные с помощью GET_HOMOLOGUES (с использованием 17 геномов для P1, 21 для P2, 21 для S1 и 5 для S2), показали U-образное распределение пангенома из четырех групп. Однако штаммы группы P1 демонстрируют асимметрию, имея в четыре раза больше отдельных видов, чем коровые гены.
https: // doi.org / 10.1371 / journal.pntd.0007270.g003
Чтобы лучше определить геномные характеристики четырех субкладов, мы сравнили различные общие характеристики, показанные на рис. 4. Члены субклада P1 часто значительно более расходятся, чем те, которые составляют другой. субклады. В целом геномы видов, принадлежащих к P1, имеют тенденцию быть больше, имеют более высокое и разбросанное содержание GC (общее для P2), несут больше генов, кодирующих тРНК (общие для P2), имеют более низкий коэффициент кодирования и большее количество псевдогенов. .Для некоторых исследованных особенностей субклад P1 имел рассеянное распределение. Другие исследования уже отметили наличие подгрупп видов и необычные расхождения в некоторых геномных характеристиках [11, 15]. Чтобы проверить, можно ли это увидеть в нашем анализе, мы разделили субклад P1 на две группы, причем одна группа — это виды, которые разошлись после определенного узла эволюции (узел 1 на рис. 1, который разделяет виды, часто участвующие в инфекциях). Обе группы существенно различаются по GC%, коэффициенту кодирования и процентному содержанию псевдогенов.
Рис. 4.
Распределение (A) общей длины, (B) GC%, (C) количества генов тРНК, (D) количества CDS, (E)% кодирования и (F)% псевдогенов (значения в log) в четырех основных субкладах. Точки, представляющие специфичные для генома значения видов, которые разошлись после узла 1 на рис. 1 ( L . interrogans , L . kirschneri , L . noguchii , L . santarosai , L . майотенз , L . боргпетерсении , L . alexanderi и L . weilii ) красного цвета. Знак «*» представляет уровень значимости между различными группами: * P ≤ 0,05, ** P ≤ 0,01, *** P ≤ 0,001 и **** P ≤ 0,0001. Уровень значимости между двумя группами P1, разделенными узлом 1, представлен одним и тем же кодом, но для ясности обозначен символом «a».
https://doi.org/10.1371 / journal.pntd.0007270.g004
Для дальнейшей характеристики субкладов CDS различных геномов были сгруппированы по функциональным категориям для оценки потенциального обогащения в одном из них (рис. 5). В общей сложности 16 категорий, участвующих в известных функциях, значительно обогащены по крайней мере одним субкладом (p <0,05). Две группы субклада P1 также можно существенно разделить на восемь из 16 категорий. Интересно отметить, что около трети CDS не были отнесены к функциональным категориям для всех субкладов и что виды, принадлежащие к субкладу P1, имеют значительно больше неназначенных CDS.Эти белки не имеют сходства с функциональными категориями, включая COG с неизвестной функцией. Это могут быть остатки псевдогенов, неправильная аннотация, а также белки, ограниченные родом Leptospira .
Рис. 5. Распределение прогнозируемых CDS по функциональным категориям (%).
Точки, представляющие специфичные для генома значения видов, которые разошлись после узла 1 на рис. 1 ( L . запросов , L . kirschneri , L . ногучи , L . santarosai , L . майотенз , L . боргпетерсении , L . alexanderi и L . weilii ) красного цвета. Знак «*» представляет уровень значимости между различными группами: * P ≤ 0,05, ** P ≤ 0,01, *** P ≤ 0,001 и **** P ≤ 0,0001. Уровень значимости между двумя патогенными группами представлен одним и тем же кодом, но для ясности обозначен символом «а».Показаны только функциональные категории, показывающие значительную разницу.
https://doi.org/10.1371/journal.pntd.0007270.g005
Наконец, мы исследовали количество липопротеинов и распределение известных факторов вирулентности обновленного рода (рис. 6 и 7). Интересно, что липопротеины, которые являются мембранными белками, кодируются меньшим числом генов в субкладе P1 по сравнению с другими субкладами; это особенно верно для видов, которые разошлись после узла 1 (наиболее вирулентные виды) внутри субклада P1 ( L . опросов , L . Киршнери , L . ногучи , L . santarosai , L . майотенз , L . боргпетерсении , L . alexanderi и L . weilii ) (рис.6). Напротив, как и ожидалось, можно наблюдать градиент в репертуаре генов, кодирующих белки, которые, как известно, участвуют в вирулентности (рис. 7A). Виды субкладов P1 и P2, имеющие наибольшее количество генов, кодирующих факторы вирулентности, и S1 и S2, имеющие наименьшее количество генов.Однако интересно отметить, что распределение гена, кодирующего каталазу KatE (LA1859), который является важным фактором вирулентности в модели на животных [38], является более гетерогенным, чем предполагалось ранее [10], поскольку можно с уверенностью определить найти гомологичную копию в геномах некоторых штаммов, принадлежащих субкладам P2, S1 и S2. Известно, что несколько доменов PFAM связаны с белками, участвующими в вирулентности Leptospira [10]. Как и ожидалось, оказалось возможным найти гораздо большее количество этих доменов у видов P1, более конкретно у видов, которые разошлись после узла 1 с высоким уровнем патогенности у людей (Рис. 7B).
Рис. 6. Распределение генов, кодирующих липопротеины.
Знак «*» представляет уровень значимости между различными группами: * P ≤ 0,05, ** P ≤ 0,01, *** P ≤ 0,001 и **** P ≤ 0,0001. Уровень значимости между двумя патогенными группами P1 (до и после узла 1) представлен одним и тем же кодом, но для ясности обозначен символом «a». Точки, представляющие специфичные для генома значения видов, которые разошлись после узла 1 на рис.1 ( L. interrogans, L.kirschneri, L. noguchii, L. santarosai, L. mayottensis, L. borgpetersenii, L. alexanderi и L. weilii ) отмечены красным.
https://doi.org/10.1371/journal.pntd.0007270.g006
Рис.
