Как разморозить страницу ВК
АВТОМАТИЗАЦИЯ ФБ
Заморозка страницы Вконтакте — частое явление, с которым сталкиваются пользователи социальной сети. Для арбитражников, которые льют трафик через этот источник, блокировка ВК означает, что они не смогут получать прибыль и продвигать товары через свой профиль. Команда diasp.pro расскажет, как разморозить страницу в ВК, а также рассмотрит все способы, с помощью которых, можно восстановить аккаунт.
Содержание
Почему ВКонтакте использует заморозкиЧаще всего, заморозки применяют, чтобы обезопасить данные пользователей от злоумышленников и защитить от возможных взломов или спам-рассылок. Также, получить бан можно за нарушение правил социальной сети. Рассмотрим основные причины, по которым модераторы могут блокировать вашу страницу:
- Взлом аккаунта или вход с другого устройства.

Автоматические алгоритмы ВКонтакте 24/7 следят за вашей активностью в Сети. При попытке взлома или входа с подозрительного устройства, система самостоятельно снимает заморозку. Такие действия позволяют обезопасить людей от кражи личных данных или от совершения незаконных действий через аккаунт. Преступники используют различные способы, чтобы получить доступ к личным данным. Они могут заразить телефон вирусом, применить специальные программы или просто подобрать логин и пароль наугад.
- Жалобы от пользователей.
Модераторы активно следят за порядком, поэтому могут заблокировать страничку вручную, если на вас было отправлено много жалоб. Причины для заморозки страницы ВК разные, однако, все они связаны с нарушением правил социальной сети. Пользователи могут пожаловаться на профиль, если вы:
- Отправляете спам или оскорбления;
- Пропагандируете наркотики или алкоголь;
- Рекламируете запрещенные товары;
- Выставляете в профиле эротический контент;
- По мнению ваших друзей или знакомых, стали жертвой мошенников;
- Создали клон известной личности.
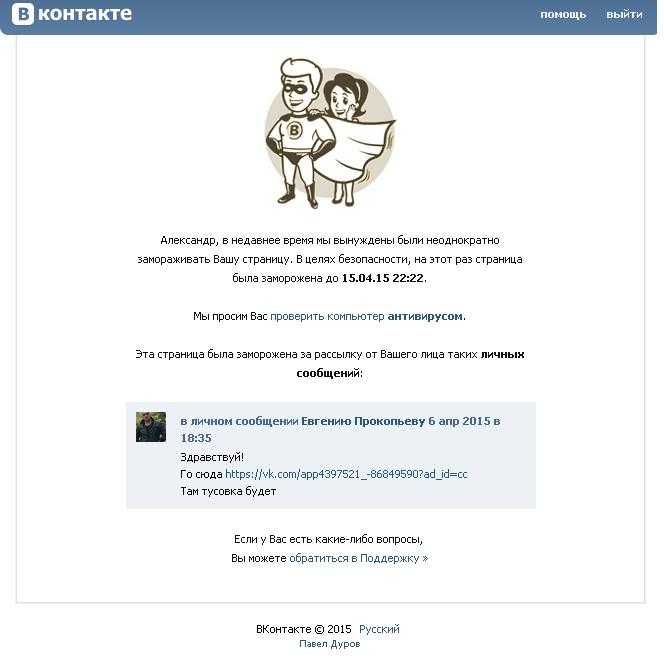
- Автоматическая блокировка.
Алгоритмы социальной сети могут заморозить учетную запись автоматически, если заметят подозрительную активность в профиле. Это может быть накрутка подписчиков и лайков, одинаковые сообщения, спам. Также подозрения вызывают массовые рассылки и приглашения в группы ВК.
Можно ли разморозить профиль ВКонтакте? Этот вопрос часто волнует пользователей мессенджера. Снять блокировку можно, но не во всех случаях. Например, если вас взломали мошенники, то аккаунт легко восстановить, как только вы подтвердите свою личность.
Если вы получили автоматический бан, то разморозить профиль будет сложнее. В таком случае,срок блокировки продолжается от нескольких часов до нескольких дней. Тем не менее, вы можете ускорить процесс и разморозить профиль, если обратитесь в поддержку и договоритесь о досрочной разморозке.
Но если вас заблокировали из-за жалоб, то разморозить страницу досрочно не получится. В таких случаях, вы не будете знать сколько продлится заморозка, пока не обратитесь в поддержку. А если вы часто нарушаете правила и получаете не первое замечание, то модераторы могут навсегда забанить ваш аккаунт и тогда отменить блокировку не получится.
В таких случаях, вы не будете знать сколько продлится заморозка, пока не обратитесь в поддержку. А если вы часто нарушаете правила и получаете не первое замечание, то модераторы могут навсегда забанить ваш аккаунт и тогда отменить блокировку не получится.
Если вы стали жертвой мошенников и на вашем аккаунте была совершена попытка взлома, то инструкция, как разморозить страницу ВКонтакте появится на экране входа.
Чтобы вернуть права на пользование, введите номер телефона, использовавшийся при регистрации, на который придет код для сброса пароля. После этого, придумайте новый пароль и сохраните изменения в настройках.
Как обезопасить аккаунт ВК от мошенниковЕсли вас взломали один раз, то это не значит, что других попыток не будет и больше не нужно следить за безопасностью данных. Приложение имеет дополнительную систему защиты — подтверждение входа или двухфакторную аутентификацию.
Подтверждение входа (двухфакторная аутентификация) — это функция, которая позволяет усилить защиту аккаунта за счет установки дополнительного кода доступа. При попытке входа с другого устройства, добавочный код будет отправлен на устройство владельца для подтверждения личности.
Таким образом, преступники не смогут зайти на вашу страничку, не зная дополнительный код, даже если подобрали логин и пароль. Включить функцию можно в настройках приложения.
Как снять заморозку, если вас забанили за нарушение правилЕсли вы нарушили правила социальной сети, то зайти в профиль и пользоваться им сразу не получится. При попытке входа, на экране появится сообщение, в котором будет указано время, через сколько вам разморозят страницу. Чтобы разморозить досрочно, нужно писать в поддержку и договариваться об условиях разморозки. Не факт, что менеджеры одобрят заявку и вернут вам доступ, если вы систематически нарушали правила.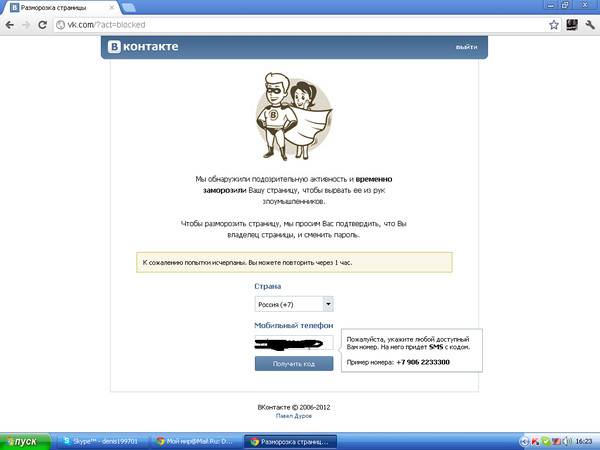
Несмотря на то, что ВК может навсегда заблокировать ваш профиль, можно попробовать написать в техподдержку и уговорить разблокировать вас. Для этого можно использовать следующие варианты:
- Отправить письмо в чат.
Не нужно проявлять агрессию, стоит спросить максимально вежливо о причинах блокировки. Также не пытайтесь обмануть поддержку в том, что вас заблокировали ошибочно, т.к. система отслеживает все действия пользователей внутри приложения. Обязательно скажите, что вы сожалеете об ошибке и подобных ситуаций больше не повторится. Однако, рассчитывать на гарантированную разморозку все же не стоит.
- Предоставить документы и фото.
Следующий вариант, как вы можете повлиять на решение — предоставить подтверждающие документы. Для этого, сделайте свою фотографию так, чтобы на фото были видны и вы, и ваша заявка о разморозке. Также нужно приложить паспорт в электронном виде в цвете в хорошем качестве.
Если фото и документы будут приняты, то шансы на разморозку повышаются. В этом случае, вас могут перенаправить на форму для восстановления сразу или через какое-то время. - Написать на электронную почту поддержки.
Самый сложный и долгий способ, который может потребоваться, если замороженный профиль регистрировался не вами. В таком случае, модераторы могут отклонить документы и фото. Последний шанс — обращаться в поддержку ВК через почту и описывать проблему там. Однако, рассчитывать на быстрое решение проблемы не стоит. Обычно, время ожидания составляет несколько дней, а весь процесс может затянуться на неделю или месяц. Если разблокировать аккаунт не получилось даже таким способом, создавайте новый профиль — ваша страница заморожена навсегда.
 Если фото и документы будут приняты, то шансы на разморозку повышаются. В этом случае, вас могут перенаправить на форму для восстановления сразу или через какое-то время.
Если фото и документы будут приняты, то шансы на разморозку повышаются. В этом случае, вас могут перенаправить на форму для восстановления сразу или через какое-то время.Заключение
Заморозка страницы ВКонтакте — не редкость. Пользователи могут получить временный бан за незначительные нарушения правил социальной сети или при попытке взлома мошенниками.
Разморозить страницу ВК, достаточно, просто. При попытке взлома, восстановить профиль получится, как только вы подтвердите свою личность и поменяете пароль. За такие нарушения, как накрутка подписчиков или рассылка спама, модераторы могут выдать временный бан, который можно досрочно снять через поддержку. Однако, если вас забанили навсегда, по причине частых жалоб от пользователей — снять блокировку не получится, и нужно создавать новый профиль.
При попытке взлома, восстановить профиль получится, как только вы подтвердите свою личность и поменяете пароль. За такие нарушения, как накрутка подписчиков или рассылка спама, модераторы могут выдать временный бан, который можно досрочно снять через поддержку. Однако, если вас забанили навсегда, по причине частых жалоб от пользователей — снять блокировку не получится, и нужно создавать новый профиль.
Удачи и профита!
Почему ВК замораживает профиль?
Как разморозить страницу ВКонтакте
Как обезопасить аккаунт от заморозки?
Что нельзя делать ВКонтакте, чтобы не получить бан?
false
Регистрация
Пароль
Подтвердите пароль
Согласие на обработку персональных данных. Подробнее
Вы уверены что хотите выйти?
ДаКак разблокировать страницу в ВК: если заблокировали на время
Содержание
- 1 Причины, по которым страницу могут заблокировать
- 2 Советы по разблокировке страницы ВК
- 2. 1 Вход по номеру телефона
- 2.2 Вход в аккаунт ВК, если номер телефона изменен
- 2.3 Можно ли восстановить страницу, если ее заблокировали навсегда
- 2.
- 3 Как уберечь аккаунт от взлома
- 4 Видео по теме
 1 Вход по номеру телефона
1 Вход по номеру телефонаВКонтакте — популярная социальная сеть, которой пользуется огромное количество людей. Там можно общаться с друзьями, слушать музыку, играть в игры, смотреть видео и многое другое. Случается, что из-за повышенной активности, аккаунт могут заблокировать. Это вызывает недоумение со стороны пользователя. Он начинает искать способы как разблокировать свою страницу в ВК.
Причины, по которым страницу могут заблокировать
Профиль в ВК есть у многих людей, начиная от детей и заканчивая продвинутыми пенсионерами. Это предоставляет аферистам широкие возможности: чем больше людей пользуется социальной сетью, тем больше страниц можно взломать. Бывает, что мошенники взламывают чужие аккаунты с целью обогатиться и просят у друзей владельца профиля деньги.
Когда ВК видит подозрительную активность, страница тут же блокируется. Мошенничество не единственная причина, по которой происходит блокировка. Также активность самого юзера может вызвать подозрения, вследствие чего профиль будет заморожен на неопределенное время. На страницу также могут подать жалобу другие пользователи. Эти причины немного размытые, но всё же существует несколько официальных. Если заблокировали страницу в ВКонтакте, нужно сначала узнать точную причину, чтобы действовать правильно. Итак, страница ВКонтакте может быть заблокирована из-за:
- Ошибки при вводе собственных данных. ВК даёт пользователям несколько попыток для ввода данных, чтобы зайти на свой профиль. Если человек раз за разом использует неверный логин или пароль, профиль может быть заблокирован. Причина максимально проста: социальная сеть думает, что в аккаунт хотят зайти злоумышленники, поэтому использует блокировку, чтобы избежать взлома.
- Жалоб других пользователей. Они чаще всего жалуются на спам, исходящий от страницы. Профиль будет временно заблокирован, пока администрация социальной сети не выяснит причины подозрительной рассылки.
- Повышенного лимита на заявки и приглашения. Профиль в Контакте может быть заморожен из-за того, что юзер отправил слишком много заявок в друзья или сообщества. Такая активность также расценивается как спам и за это могут запросто заблокировать страницу.
- Взлома аккаунта мошенниками. Если какой-то чужой человек зашел в профиль юзера, то он автоматически утрачивает к нему доступ. Чтобы разблокировать страницу в ВК, следует обратиться к администрации соцсети.
- Противоречащего правилам сообщества контента в профиле. Есть определенные правила, которыми оговаривается, какую информацию нельзя размещать на своей странице ВКонтакте. К ней относятся посты, пропагандирующие терроризм, наркотики и так далее. Обычно модераторы ВК удаляют только противоречащие правилам записи, но часто происходит и блокировка аккаунта человека, который их размещал.
 Профиль будет временно заблокирован, пока администрация социальной сети не выяснит причины подозрительной рассылки.
Профиль будет временно заблокирован, пока администрация социальной сети не выяснит причины подозрительной рассылки.Советы по разблокировке страницы ВК
Существует несколько способов, с помощью которых можно решить вопрос, что делать пользователю, если его страницу в ВК заблокировали. Они действуют и на телефонах (iOS, Android), и на компьютерах.
Они действуют и на телефонах (iOS, Android), и на компьютерах.
Вход по номеру телефона
Этот способ действует только в том случае, когда профиль забанили в первый раз. Чтобы разблокировать его, нужно зайти в приложение и ввести свой номер телефона в отдельное поле. После этого нажать на «Ввод». На экране появится надпись, что на телефон отправлен код подтверждения.
Код нужно ввести в специальную графу в приложении и отправить его. После подтверждения личности, ВК выбросит юзера на страницу, где ему надо будет придумать новый пароль для входа в аккаунт. Если аккаунт в ВК заблокировали из-за взлома мошенниками, то новый пароль должен быть намного надежнее предыдущего, чтобы снизить риск вскрытия снова.
Вход в аккаунт ВК, если номер телефона изменен
Иногда юзерам приходится искать ответ на вопрос, что следует делать, если заблокировали страницу ВКонтакте, зарегистрированную по номеру телефона, которого больше не существует, и нет возможности получить к нему доступ. В этом случае не стоит отчаиваться, поскольку разблокировка профиля возможна и без номера телефона.
В этом случае не стоит отчаиваться, поскольку разблокировка профиля возможна и без номера телефона.
Но, чтобы разблокировать ВК в этом случае, придется потрудиться. Сначала необходимо привязать аккаунт к другому телефону, который ранее не применялся. Если у пользователя есть вторая СИМ-карта, то он может смело воспользоваться этим способом. В противном случае следует искать другой метод.
Чтобы разблокировать профиль, нужно зайти в приложение ВКонтакте и в специальном окне ввести свой номер телефона. Далее юзеру необходимо прикрепить фото любого его документа, чтобы подтвердить, что страница действительно принадлежит ему. После этого в свободном поле нужно объяснить причину, по которой доступ к аккаунту был утерян. Если его взломали, то об этом также стоит написать.
Можно ли восстановить страницу, если ее заблокировали навсегда
ВКонтакте, как и любая другая социальная сеть хочет иметь как можно больше пользователей, поэтому блокировка аккаунтов навсегда осуществляется здесь только по очень серьезной причине. Если это произошло, значит, пользователь нарушил правила сообщества. Юзерам, которых интересует, что надо делать, если тебя заблокировали в ВК навсегда, следует знать, что шанс вернуть профиль очень мал. Тем не менее, можно попробовать.
Если это произошло, значит, пользователь нарушил правила сообщества. Юзерам, которых интересует, что надо делать, если тебя заблокировали в ВК навсегда, следует знать, что шанс вернуть профиль очень мал. Тем не менее, можно попробовать.
Самым верным способом является разговор со службой поддержки. Для этого следует войти в ВК с телефона или компьютера и найти кнопку для обращения в техподдержку. Когда откроется окно с пустым полем, нужно будет заполнить его, описав свою проблему максимально подробно.
Чтобы увеличить шанс вернуть аккаунт, нужно придерживаться трёх основных правил при общении с сотрудниками технической поддержки:
- Рассказывать о своей проблеме надо чётко и подробно. Если пользователь всё хорошо объяснит, то техподдержка поймёт и постарается решить проблему.
- Следует предоставить доказательства того, что аккаунт был заблокирован беспричинно. Этот вариант подходит для тех, кто действительно не делал ничего противоречащего правилам сообщества. Если же юзер нарушил правила, то следующий пункт для него.
- Тем, кто нарушил правила ВКонтакте и их аккаунт был заморожен, следует признать свою вину и попросить у технической поддержки прощения за свой проступок.
 Если же юзер нарушил правила, то следующий пункт для него.
Если же юзер нарушил правила, то следующий пункт для него.Также есть список того, чего не стоит делать при общении с сотрудниками:
- Ни в коем случае нельзя грубить и угрожать. Техподдержка работает исключительно в интересах пользователей. Если юзер действительно хочет, чтобы проблема была решена, ему следует быть культурным и вежливым.
- Нельзя оскорблять других пользователей социальной сети, которые, по мнению юзера, виноваты в блокировке аккаунта.
К сожалению, не всегда сотрудники технической поддержки могут помочь пользователям в решении данной проблемы. Это может сработать, если профиль был заблокирован без весомой причины или по ошибке. Если же человек сам виноват в этом (распространял спам, выкладывал неприемлемые видео о наркотиках и так далее), то тут вряд ли чем-то можно будет сделать.
Как уберечь аккаунт от взлома
Ни один способ не даёт стопроцентной гарантии того, что профиль не взломают. Тем не менее, если предпринять определенные действия, риск взлома сильно снизится. Лучше всего от мошенников защищает надёжный пароль. Поэтому следует придумать длинную комбинацию из букв, цифр и символов. Это снижает шанс взлома аккаунта, поскольку подобрать длинный и сложный код весьма затруднительно.
К сожалению, даже самый сложный пароль не обеспечивает абсолютной гарантии. Ведь аферисты пользуются нелегальными приложениями и программами для вычисления кода. Несмотря на это, составить надежную комбинацию стоит хотя бы для большей уверенности и собственного спокойствия.
Не нужно расстраиваться и паниковать, если заблокирован аккаунт в ВКонтакте, что делать при этом было рассказано в статье. Чтобы снизить риск блокировки, следует исключить распространение спама и контента, запрещенного в социальной сети, на своей странице. Также надо стараться не подавать слишком много заявок в друзья и не вступать сразу в много сообществ, поскольку ВК расценивает это как подозрительную активность. ВКонтакте никогда не блокирует аккаунты без причины. Если чётко соблюдать правила, пользование социальной сетью будет приятным.
ВКонтакте никогда не блокирует аккаунты без причины. Если чётко соблюдать правила, пользование социальной сетью будет приятным.
Видео по теме
Эффективный способ, как разблокировать страницу в ВК?
17:00, 29 октября 2018
Как работать
Автор: Amelie
Сегодня ВКонтакте активно используется в коммерческих целях. Люди все чаще прибегают к различным запрещенным приемам для ускорения процесса продвижения, что зачастую приводит к бану. У каждого, кто попадал в подобную ситуацию, возникает вопрос, как разблокировать страницу ВК? И сегодня я дам на него ответ.
В каких случаях блокируют страницу ВК?
Как и любого другого сайта, у vk. com есть свои правила, нарушение которых и приводит к заморозке или блокировке аккаунта. В большинстве случаев, особенно если нарушение конкретного профиля зафиксировано впервые, страницу именно замораживают на определенный срок, а не банят навсегда. В таких ситуациях разблокировать ее куда проще. Если же в сообщении от администрации говориться о бане навсегда, тогда, увы, шансы на разбан сводятся к нулю.
com есть свои правила, нарушение которых и приводит к заморозке или блокировке аккаунта. В большинстве случаев, особенно если нарушение конкретного профиля зафиксировано впервые, страницу именно замораживают на определенный срок, а не банят навсегда. В таких ситуациях разблокировать ее куда проще. Если же в сообщении от администрации говориться о бане навсегда, тогда, увы, шансы на разбан сводятся к нулю.
Итак, вот список наиболее популярных причин блокировки:
- Спам (рассылка личных сообщений или комментариев с целью рекламирования продукта). Смотрите о лимитах на отправку сообщений, в материале по ссылке.
- Публикация запрещенных видео или фотоматериалов.
- Массовая рассылка приглашений в группу и в друзья.
- Размещение вредоносных ссылок.
- Использование сервисов, искусственно увеличивающих счетчики активности (накрутка).
- Упоминание таких сервисов или их рекламирование.
- Оскорбления и т.д.
Это лишь наиболее распространенные причины. В большинстве случаев аккаунт будет заморожен. А вот если нарушение носит более серьезный характер (например, призывы к насилию, терроризму, экстремизму и т.д.), можно получить перманентный бан. Советую ознакомиться.
В большинстве случаев аккаунт будет заморожен. А вот если нарушение носит более серьезный характер (например, призывы к насилию, терроризму, экстремизму и т.д.), можно получить перманентный бан. Советую ознакомиться.
Как разблокировать?
Что делать, если страницу заблокировали? Правильно, необходимо восстановить к ней доступ.
Чтобы разблокировать страницу ВК, необходимо открыть сайт на ПК (именно десктопную версию сайта, а не мобильную). Дальнейшие действия зависят от причины и вида блокировки.
Последовательность разблокирования профиля следующая:
- Авторизоваться под своим логином и паролем (чтобы появилось сообщение о бане).
- Если указано время заморозки, дождаться ее окончания.
- Ввести номер телефона.
- Далее полученный в СМС код ввести в появившееся поле.
- Указать новый пароль.
- Прикрепить скан или фото документов, подтверждающих личность.
- Прикрепить свое фото на фоне сообщения о блокировке.

Последние пункты требуются не всегда. Если вас впервые заморозили, без указания срока, то для восстановления доступа достаточно получить СМС-сообщение, ввести код и новый пароль.
Если вас заблокировали не в первый раз, то здесь уже администрация ВК может потребовать скан паспорта или вашу фотографию на фоне текущей сессии. Это куда проблематичнее, чем подтверждение через СМС. И если со сканом паспорта все намного проще, то личное фото может осложнить процесс разбана страницы. Фото должно быть четким, иметь хорошее освещение, чтобы ваше лицо было отчетливо видно. При этом на фотографии также должна быть видна и текущая сессия (сообщение от администрации сайта). Учитывая, что большинство камер не может одновременно сфокусировать и картинку экрана и то, что находится за его пределами, у большинства пользователей возникают трудности.
Поэтому, если вас неоднократно наказывали за нарушение правил сайта, лучше приостановить всю подозрительную деятельность или использовать другой аккаунт.
Вам будет интересно: “Бережёного бог бережёт: защитим от покушений страницу ВК“.
Как разблокировать страницу без телефона?
Бывают ситуации, когда у владельца замороженной страницы по тем или иным причинам нет возможности использовать свой телефонный номер. Что делать в таких ситуациях? – Восстанавливать страницу через функцию «Забыли пароль».
Как разблокировать страницу ВК без номера телефона:
- Откройте десктопную Версию сайта ВК.
- Нажмите «Забыли пароль».
- Далее кликните по ссылке «Нажмите сюда».
- Введите ссылку своего профиля.
- Введите новый и старый номер, старый e-mail и пароль, привязанные к аккаунту.
- Отправьте запрос.
Далее ваш запрос будет рассмотрен администрацией сайта, и они уже объяснят ваши дальнейшие действия.
Итог
Если ваш профиль заморожен, то вы всегда сможете восстановить к нему доступ. Если бан получен навсегда, то проще завести новый аккаунт. Сегодня я рассказал вам о том, как разблокировать страницу в ВК.
А о том, как разблокировать свой номер в Телеграм, есть статья на нашем сайте.
Читайте далее:
Click to rate this post!
[Total: 1 Average: 5]
«Ярче!» — сеть супермаркетов.
Сеть «Ярче!»
«Ярче!» доставляет
Заказывайте свежие продукты
в интернет-магазине yarcheplus.ru, экономьте время и деньги!
Новый день — новая партия свежих продуктов!
В наших супермаркетах нет складов, поэтому продукты повседневного спроса в сеть «Ярче!» и интернет-магазин доставляют ежедневно.
У нас нет от вас секретов!
Мы подробно рассказываем о том,
как производят продукты для наших
супермаркетов. Давайте вместе
взглянем на фабрики!
Все видео image/svg+xml Посмотреть вакансииДля вас в Сибири и Московской области работает более 650 супермаркетов «Ярче!».
Каждый день в них совершают покупки не менее 1 000 000 человек
С пекарней Без пекарни
Регион
Алтайский край
Владимирская область
Ивановская область
Кемеровская область
Красноярский край
Московская область
Новосибирская область
Омская область
Республика Мордовия
Рязанская область
Тверская область
Томская область
Тульская область
Ярославская область
Населенный пункт
Всег. Анжеро-Судженск
г. Белово
п. Березовский
г. Гурьевск
п. Елыкаево
п. Ижморский
п. Калтан
г. Кемерово
г. Киселевск
п. Краснобродский
г. Ленинск-Кузнецкий
г. Мариинск
г. Междуреченск
г. Мыски
г. Новокузнецк
г. Осинники
п. Поломошное
г. Полысаево
г. Прокопьевск
п. Промышленная
п. Сосновка
г. Тайга
п. Топки
г. Юрга
п. Яшкино
п. Яя
- г. Анжеро-Судженск
- ул. 50 лет Октября, 1а
- ул. им.137 Отдельной Стрелковой Бригады, 4
- ул. Коминтерна, 24
- ул. Куйбышева, 61
- ул. Перовской, 19
- ул. Челинская, 8
- г. Белово
- 3-й микрорайон, 14г
- пгт. Бачатский, ул. Шевцовой, 58
- пгт. Грамотеино, ул. Колмогоровская, 34
- ул. Доватора, 8
- ул. Киевская, 27
- ул. Ленина, 65
- ул. Октябрьская, 31
- ул. Светлая, 1а
- ул. Советская, д. 37а
- ул. Юбилейная, 11а
- п. Березовский
- б-р Молодёжный, 8
- пр. Ленина, 15
- ул. Волкова, 1б
- ул. Карбышева, 5
- г. Гурьевск
- ул. Ленина, 85
- п. Елыкаево
- ул. Советская, 75
- п. Ижморский
- ул. Ленинская, 68а
- п. Калтан
- пр. Мира, 40
- ул. Дзержинского, 51
- ул. Комсомольская, 55
- г. Кемерово
- б-р Осенний, 1
- б-р Строителей, 29
- б-р Строителей, 42г
- пр. Комсомольский, 11б
- пр. Комсомольский, 34
- пр. Комсомольский, 45
- пр. Ленина, 136
- пр. Ленина, 26
- пр. Ленина, 51б
- пр. Ленина, 63
- пр. Ленина, 82
- пр. Ленинградский, 22
- пр. Молодежный, 15б
- пр. Октябрьский, 10
- пр. Октябрьский, 68
- пр. Октябрьский, 9
- пр. Притомский, 31/1
- пр. Притомский, 7/3
- пр. Шахтеров, 39
- пр. Шахтеров, 50
- пр. Шахтеров, 57
- ул. 4-я Цветочная, 4
- ул. 50 лет Октября, 21
- ул. 9 Января, 6
- ул. Базовая, 14
- ул. Белозерная, 15
- ул. Белозерная, 44
- ул. Благовещенская, 53
- ул. Варяжская, 22а
- ул. В.Волошиной, 28
- ул. Ворошилова, 14
- ул. Двужильного, 10
- ул. Дзержинского, 4
- ул. Коломейцева, 10
- ул. Космическая, 22
- ул. Красноармейская, 95
- ул. Леонова, 22
- ул. Леонова, 4
- ул. Мичурина, 43
- ул. Новогодняя, 16
- ул. Севастопольская, 3
- ул. Серебряный бор, 15А
- ул. Сибиряков-Гвардейцев, 11
- ул. Терешковой, 38
- ул. Тухачевского, 16
- ул.Тухачевского, 31
- ул. Халтурина, 37а
- ул. Цимлянская, 2
- г. Киселевск
- ул. 50 лет города, 38
- ул. 50 лет Октября, 43
- ул. Ленина, 39
- ул. Черноморская, 23а
- п. Краснобродский
- ул. Комсомольская, 11
- г. Ленинск-Кузнецкий
- пр. Кирова, 69
- пр. Ленина, 53/1
- пр. Ленина 71а
- пр. Ленина, 90Д
- пр. Текстильщиков, 3
- пр. Текстильщиков, 4/4
- пр. Химиков, 10/1
- ул. 7-ое ноября, 32
- ул. Горького, 19
- ул. Садовая, 78
- ул. Шишлянникова, 14
- г. Мариинск
- ул. Юбилейная, 28г
- г. Междуреченск
- пр. 50 лет Комсомола, 40
- пр. Строителей, 19
- пр. Шахтеров, 63
- ул. Брянская, 12
- ул.Интернациональная, 43
- ул. Коммунистический, 36
- ул. Кузнецкая, 24
- ул. Строителей, 46
- г. Мыски
- ул. 17 Квартал, д.18
- ул. Первомайская, 30
- ул. Советская, 30
- г. Новокузнецк
- микрорайон 13, 12
- пр. Бардина, 2
- пр. Бардина, 42
- пр. Дружбы, 20а
- пр. Дружбы, 3
- пр. Дружбы, 35б
- пр. Металлургов, 37
- пр. Октябрьский, 13
- пр. Октябрьский, 31
- пр. Октябрьский, 47
- пр. Октябрьский, 62а
- пр. Строителей, 71
- пр. Шахтеров, 34а
- пр. Шахтеров, 5а
- ул. 40 лет ВЛКСМ, 3г
- ул. 40 лет ВЛКСМ, 6
- ул. 40 лет ВЛКСМ, 86
- ул. 40 лет Победы, 3а
- ул.Архитекторов, 14а
- ул. Белана, 5
- ул. Веры Соломиной, 9
- ул. Горьковская, 31
- ул. Горьковская, 35а, корп.1
- ул. Грдины, 35
- ул. Грдины 7
- ул. День шахтера, 8
- ул. Ермакова, 5
- ул. Запорожская, 21
- ул. Запорожская, 81
- ул. Зорге, 46а
- ул. Кирова, 23а
- ул. Кирова, 51а
- ул. Кирова, 97
- ул. Ленина, 40
- ул. Ленина, 49
- ул. Ленина, 87
- ул. Ленина, 9
- ул. Мурманская, 49
- ул. Новобайдаевская, 7б
- ул. Новоселов, 29
- ул.Новоселов, 67
- ул. Ноградская, 4а
- ул. Обнорского, 4
- ул. Прежевальского, 26
- ул. Пролетарская, 148
- ул. Разведчиков, 52
- ул. Сеченова, 10а
- ул.Тольятти, 27
- ул. Тореза, 20
- ул. Тореза, 52
- ул. Тореза, 64
- ул. Тореза, 93
- ул. Транспортная, 43
- ул. Транспортная, 77
- ул. Чернышова, 4а
- ул. Ярославская, 12
- г. Осинники
- ул. Победы, 24
- ул. Революции, 5
- п. Поломошное
- ул. Бениваленского, 10а
- г. Полысаево
- п. Красногорский, ул. Магистральная, 19
- ул. Космонавтов, 100
- г. Прокопьевск
- микрорайон 10, 18
- мкр. 10-й, 5б
- пр. Ленина, 25
- ул. Гагарина, 32
- ул. Институтская, 104
- ул. Комсомольская, 1
- ул. Кучина, 2
- ул. Оренбургская, 8
- ул. Союзная, 56
- ул. Строителей, 7а
- ул. Шишкина, 12
- п. Промышленная
- ул. Некрасова, 1
- п. Сосновка
- ул. Калинина, 61
- г. Тайга
- ул. Советская, 86
- п. Топки
- микрорайон Солнечный, 11а
- ул. Революции, 113
- г. Юрга
- б-р. Металлургов, 3а
- пр. Кузбасский, 16
- пр. Победы, 41а
- ул. Вокзальная, 58
- ул. Леонова, 6А
- ул. Максименко, 2
- ул. Машиностроителей, 12
- ул. Мира, 13
- ул. Павлова, 16
- п. Яшкино
- ул. Молодежная, 1а
- п. Яя
- ул. Школьная, 1
Список супермаркетов «Ярче!
Военная операция на Украине. Главное — РБК
США анонсировали новый пакет военной помощи Киеву, Украина сооружает новую линию противотанковых рвов на границе с Белоруссией, США ввели санкции еще против шести россиян. Главное о последних событиях — в материале РБК *
Фото: Валентин Огиренко / Reuters
Главное о спецоперации на Украине к 21:55 мск
-
В Севастополе похоронили двоих погибших в зоне военной операции военнослужащих, сообщил губернатор региона Михаил Развожаев в телеграм-канале.
Погибшим — стрелку мотострелкового отделения 126-й отдельной гвардейской бригады береговой обороны Владимиру Шабаеву и командиру штурмового подразделения отряда «БАРС» старшему лейтенанту Сергею Логвиненко — было по 57 лет. Шабаев погиб в Херсонской области, Логвиненко — в поселке Белогоровка в ЛНР, сообщил губернатор. -
США выделяют Украине новый пакет военной помощи на $400 млн, сообщил госсекретарь США Энтони Блинкен. Он будет включать боеприпасы для ракетных установок HIMARS и артиллерийских систем калибра 155-мм и 105-мм. Кроме того, США впервые предоставят Киеву мостоукладчики.
-
Украина сооружает новую линию противотанковых рвов в Ровенской области на границе с Белоруссией, сообщил глава этого региона Виталий Коваль в своем телеграм-канале.
Как отмечает «Страна.ua», ранее украинские власти расширили до 2 км пограничную полосу вдоль границы с Россией и Белоруссией. В Госпогранслужбе говорили, что данная территория будет заминирована. -
США ввели санкции против шести россиян в соответствии с «законом Магнитского». Под ограничения попали заместитель министра юстиции России Олег Свириденко, судья Басманного суда Москвы Елена Ленская, судья Хамовнического районного суда Диана Мищенко, судья Мосгорсуда Илья Козлов, участник ряда судебных процессов в статусе свидетеля обвинения, директор АНО «Независимый эксперт» Данила Михеев и следователь по особо важным делам СК Андрей Задачин. Трое из них — Ленская, Михеев и Задачин — по мнению американского Минфина, причастны к нарушению прав оппозиционера Владимира Кара-Мурзы (внесен в реестр физлиц-иноагентов).
-
Министерство юстиции Украины подало иск, чтобы изъять находящиеся в стране активы, якобы принадлежащие бизнесменам Игорю и Аркадию Ротенбергам, а также Александру Скоробогатько и Александру Пономаренко.
Об этом сообщила пресс-служба ведомства. Исковое заявление подано в рамках применения закона «О санкциях», говорится в сообщении. Украинский Минюст просит изъять в пользу государства 100% доли уставного капитала ООО «Авангард-Виларти» и 66% доли уставного капитала ООО «Инвестиционный союз «Лыбидь».
Хроника боевых действий
Поставки зарубежного оружия Украине
Какие санкции ввели против России
Экстренное обращение Владимира Путина
adv.rbc.ru
Что предшествовало военной операции
-
Мелитополь официально стал столицей Запорожской области. Соответствующий указ подписал врио главы региона Евгений Балицкий. В документе также указано, что кроме него городами областного значения стали Бердянск и Энергодар. Соответствующие решения приняты «в целях эффективного государственного управления и оказания населению Запорожской области качественных и своевременных услуг».
-
Возле города Енакиево в ДНР сбили самолет, два летчика катапультировались, сообщил мэр Роман Храменков.
Принадлежность самолета пока не установлена, к месту происшествия выдвинулись экстренные службы. -
Проект закона о повышении призывного возраста в России планируется внести в Госдуму весной. Если его успеют принять до летних каникул, то осенью он уже может вступить в силу, заявил глава оборонного комитета верхней палаты Виктор Бондарев.
-
С начала российской военной операции Сербия не экспортировала и не выдавала лицензий на экспорт оружия и военной техники ни на Украину, ни в Россию, сообщил глава сербского МИДа Ивица Дачич, передает РТРС. Он выразил уверенность в этом, поскольку МИД участвует в выдаче лицензий на экспорт оружия. При этом министр подчеркнул, что речь идет о легальных учреждениях, которые могут экспортировать оружие.
-
США с начала военной операции России на Украине выделили более $30 млрд на помощь в области безопасности — ВСУ обещали предоставить артиллерию, ракеты, стрелковое оружие, бронетехнику, радарные системы, бронежилеты, миллионы патронов и многое другое, пишет WSJ.
США выделят Украине очередной пакет военной помощи на $400 млн и объявят об этом в пятницу, 3 марта, узнал Reuters. -
Глава компании Millhouse и деловой партнер бизнесмена Романа Абрамовича Евгений Швидлер (Forbes оценивает его состояние в $1,7 млрд) решил оспорить в лондонском суде введенные против него санкции Великобритании, пишет Bloomberg. Заявление, пишет агентство, адвокаты Швидлера подали в феврале. Они считают, что Британия допустила «существенные ошибки» в своей оценке деловых отношений истца с Романом Абрамовичем.
-
Нью-йоркский Метрополитен-музей на своем сайте изменил подпись к картине «Корабль в лунном свете» художника Ивана Айвазовского (урожденный Ованнес Айвазян, 1817–1900), выставленной в его собрании. «Армянин, родился в Российской империи (ныне Украина)», — говорится в подписи к картине. Айвазовский родился в Крыму, в декабре 2021 года на странице не было подписи к картине, а художника указывали как русского.
-
Брянский мальчик Федор по поручению председателя СК Александра Бастрыкина награжден ведомственной медалью «Доблесть и отвага». «Федор мог быстро убежать, однако он не оставил младших девочек и помог им выйти из машины через заднюю дверь, а затем скомандовал бежать в лес. Вооруженные мужчины в этот момент цинично открыли стрельбу по безоружным детям. Федор получил ранение грудной клетки, однако даже в этой ситуации он не растерялся и сумел проводить девочек через лес, поймать попутную машину и доехать до безопасного места», — говорится в сообщении.
21 сентября министр обороны России Сергей Шойгу назвал объем потерь в спецоперации. По его словам, потери России составили 5937 человек. Украина с начала спецоперации потеряла более 100 тыс. человек — 61 207 погибшими и 49 368 ранеными — это половина ее первоначальной армии.
1 декабря Минобороны России сообщило, что в ходе наступления под полный контроль был взят населенный пункт Курдюмовка (к югу от Артемовска/Бахмута).
30 ноября Министерство обороны сообщило, что в ходе наступления на Донецком направлении были заняты населенные пункты Белогоровка и Перше Травня (украинские власти переименовывали его в Озаряновку). Позже в этот день Минобороны сообщило об установлении полного контроля над населенным пунктом Андреевка (находится к югу от Артемовска) в ДНР.
18 ноября Минобороны официально сообщило о занятии поселка Опытное к югу от Авдеевки.
14 ноября Минобороны России сообщило, что после упорных боев российские войска полностью заняли в Донецкой народной республике село Павловка (к югу от Угледара).
18 октября в Минобороны России сообщили, что подразделения российской армии овладели населенным пунктом Горобьевка Харьковской области (к северо-востоку от Купянска).
7 октября штаб территориальной обороны ДНР сообщил о том, что в районе Бахмута заняты населенные пункты Отрадовка, Веселая долина и Зайцево. О занятии Зайцево Минобороны России сообщало накануне.
22 сентября штаб территориальной обороны ДНР сообщил о том, что группировка войск ДНР и ЛНР при огневой поддержке ВС России заняла населенный пункт Жованка (к югу от Бахмута).
15 сентября в штабе территориальной обороны ДНР сообщили, что военнослужащие Донецкой и Луганской народных республик взяли под контроль поселок Майорск, огневую поддержку в этом оказывали российские войска. Поселок Майорск находится на северной окраине города Горловка в Донецкой области. В этой местности находился один из контрольных пунктов въезда-выезда из ДНР на территорию Украины. Штаб теробороны республики также опубликовал карту, согласно которой дальнейшее продвижение войск планируется продолжить на север — в направлении Бахмута.
14 сентября штаб территориальной обороны ДНР сообщил о занятии населенных пунктов Николаевка и Николаевка Вторая к югу от Бахмута.
10 сентября Минобороны России сообщило о проведении операции по свертыванию и переброске изюмско-балаклейской группировки войск на территорию ДНР.
7 сентября Минобороны России сообщило о занятии Кодемы (к юго-востоку от Бахмута/Артемовска). По заявлению оборонного ведомства, украинские войска наступлений за последние сутки не проводили.
6 сентября штаб территориальной обороны ДНР сообщил о занятии еще двух населенных пунктов. Речь идет о Времевке и Новополе (к западу от Великой Новоселки).
В этот же день в Минобороны России сообщили о продолжающихся попытках украинской армии атаковать на отдельных участках николаево-криворожского направления.
2 сентября министр обороны России Сергей Шойгу подтвердил, что Украина продолжает попытки вести наступление на николаево-криворожском и других направлениях. По словам министра, «противник несет значительные потери».
1 сентября Минобороны сообщило о высадках украинского десанта в районе Запорожской АЭС. Один десант на семи лодках в составе двух групп по 60 человек высадился около 06:00 мск на побережье Каховского водохранилища в 3 км северо-восточнее станции. Они были блокированы российскими силами с помощью вертолетов армейской авиации. Три человека из группы к вечеру были захвачены в плен, часть военнослужащих продолжала сопротивление, часть погибла. Вторая группа вышла из Никополя на двух самоходных баржах и попыталась высадиться около 07:00 мск в нескольких километрах от АЭС, но была, по сообщению оборонного ведомства, в результате обстрела потоплена вместе с баржами. Эта группа, предположительно, была направлена к АЭС в качестве подкрепления первому десанту.
В тот же день Минобороны сообщило о двух новых попытках наступления украинских сил: на южном направлении, в районе Николаева и под Харьковом. По сообщению оборонного ведомства, успеха украинцам добиться не удалось.
31 августа Минобороны вновь сообщило о попытках атак украинских войск на николаево-криворожском направлении. ВСУ атаковали населенные пункты Архангельское, Ольгино и Терновые Поды, атаки были отбиты, ВСУ понесли значительные потери и отброшены.
29 и 30 августа Минобороны сообщало о наступлении ВСУ на юге страны: в Херсонской и Николаевской областях, а также со стороны Кривого Рога. Российские военные отбили атаки, сообщили о разгроме наступавших частей, многочисленных потерях среди украинских войск и значительном количестве уничтоженной техники. Заявления Минобороны об украинским наступлении появились после публикации CNN о подготовке украинской стороной необходимых условий «для серьезного контрнаступления». Изначально в офисе Зеленского опровергали появлявшиеся в СМИ сообщения о планах наступления на юге страны, однако затем пресс-секретарь Южного военного командования Украины Наталья Гуменюк подтвердила их.
24 августа Минобороны отчиталось о том, сколько украинских вооружений уничтожили российские военные за полгода спецоперации. Среди них 268 самолетов, 148 вертолетов, 1803 беспилотника, 369 зенитных ракетных комплексов, 4382 танка и других боевых бронированных машин, 819 боевых машин реактивных систем залпового огня, 3339 орудий полевой артиллерии и минометов, а также 5034 единицы специальной военной автомобильной техники.
23 августа российские военные вышли к административной границе Николаевской области, сообщило Минобороны. По данным ведомства, им удалось установить контроль над территорией области площадью 12 кв. км. Министерство заявило о взятии под контроль 36 кв. км территории Херсонской области, в том числе поселка Комсомольское.
21 августа глава Чечни Рамзан Кадыров заявил об освобождении поселка Пески под Донецком; российское Минобороны еще неделю назад заявляло о полном контроле над населенным пунктом.
18 августа штаб территориальной обороны ДНР сообщил, что группировка войск донбасских республик «при огневой поддержке ВС РФ» освободила 270 населенных пунктов на территории республики, включая Зайцево и Дачу (оба находятся к северу от Горловки).
17 августа штаб территориальной обороны ДНР сообщил о контроле над населенным пунктом Вершина (к юго-западу от Артемовска/Бахмута).
14 августа Минобороны России сообщило о занятии населенного пункта Уды в Харьковской области.
13 августа Минобороны России сообщило о занятии поселка Пески (к западу от Донецка).
3 августа штаб территориальной обороны ДНР сообщил о занятии населенных пунктов Травневое и Семигорье (к югу от Артемовска/Бахмута).
29 июля штаб территориальной обороны ДНР сообщил о контроле над селами Клиновое и Покровское (к востоку от Бахмута).
26 июля помощник главы МВД ЛНР Виталий Киселев сообщил о занятии Углегорской ТЭС, которая находится напротив Светлодарска (к востоку от Бахмута/Артемовска).
В этот же день Киселев сообщил о занятии Новолуганского (к востоку от Бахмута/Артемовска).
20 июля штаб территориальной обороны ДНР сообщил о занятии села Берестовое (к северу от Соледара).
19 июля штаб территориальной обороны ДНР сообщил о занятии села Старомайорское (к югу от Великой Новоселки).
14 июля Народная милиция ЛНР сообщила о занятии населенных пунктов Стряповка и Новая Каменка (к востоку от Соледара).
14 июля штаб территориальной обороны сообщил о занятии Народной милицией ДНР при поддержке российской армии следующих населенных пунктов: Новодонецкое, Нескучное, Октябрь (к югу от Великой Новоселки), Мироновку, Красный Пахарь, Роты, Видродження, Медную Руду (к востоку от Артемовска/Бахмута) и Каменку (к северу от Авдеевки).
13 июля помощник главы МВД ЛНР Виталий Киселев заявил агентству ТАСС, что силы Народной милиции зашли в городскую черту Соледара (к югу от Лисичанска). Ранее ТАСС со ссылкой на Киселева сообщал о начале боев за Северск.
12 июля Минобороны России обвинило украинских военных в минировании мостов через реку Северский Донец в селе Николаевка (ДНР). «Они намерены их взорвать и по уже отработанному сценарию с широким освещением в украинских и западных СМИ обвинить российские вооруженные силы в якобы неизбирательных ударах по критически важным объектам транспортной инфраструктуры», — говорится в сообщении.
Тогда же, 12 июля, глава ЛНР Леонид Пасечник заявил, что подразделения Народной милиции продолжат наступление, поскольку украинские войска продолжают обстреливать территорию республики. «Подразделения Народной милиции ЛНР вынуждены будут продолжить наше движение вперед, чтобы отодвинуть противника на безопасное расстояние, чтобы имеющаяся у них техника и вооружение, боеприпасы не доставали до нашей территории», — цитирует его Луганский информационный центр.
11 июля официальный представитель Народной милиции ЛНР Иван Филипоненко сообщил о взятии силами республик Донбасса населенных пунктов Владимировка и Триполье в ДНР (к востоку от Соледара).
10 июля посол ЛНР в России Родион Мирошник сообщил, что войска продолжают наступление на город Северск в ДНР. Он также рассказал о взятии населенного пункта Григоровка.
6 июля штаб территориальной обороны ДНР сообщил о занятии населенного пункта Спорное (к востоку от Северска).
3 июля Сергей Шойгу доложил Владимиру Путину, что Вооруженные силы России совместно с подразделениями Народной милиции Луганской народной республики установили полный контроль над городом Лисичанск.
Фото: Александр Ратушняк / EPA / ТАСС
2 июля Минобороны сообщило, что в результате успешных наступательных действий войска группировки «Центр» под командованием генерал-полковника Александра Лапина овладели населенными пунктами Верхнекаменка, Золотаревка, Белогоровка. Помимо этого были заняты населенные пункты Новодружеск, Малорязанцево и Белая Гора.
1 июля Минобороны сообщило о продвижении Вооруженных сил России и формирований ЛНР и ДНР под Лисичанском и их выходе непосредственно к городу. Были взяты под контроль Лисичанский нефтеперерабатывающий завод, шахта «Матросская», желатиновый завод, а также поселок Тополевка.
В этот же день представитель Народной милиции ЛНР сообщил о занятии сел Приволье и Шипиловка (к западу от Лисичанска).
Днем 30 июня в штабе территориальной обороны ДНР заявили о занятии села Сидорово (расположено к югу от Святогорска).
Утром 30 июня Минобороны сообщило о выводе «в качестве шага доброй воли» гарнизона с острова Змеиный в Черном море.
27 июня помощник главы МВД ЛНР Виталий Киселев заявил в эфире телеканала «Россия 24», что республиканские силы и российские военные завершили зачистку Волчеяровки (поселок к юго-западу от Лисичанска).
26 июня Минобороны России сообщило о занятии Северодонецка и расположенных рядом населенных пунктов (в том числе Вороново и Боровское).
24 июня Минобороны России сообщило, что в результате успешных наступательных действий группировки российских войск на луганском направлении за пять дней были освобождены населенные пункты Лоскутовка, Подлесное, Мирная Долина, Щебкарьер, Врубовка, Нырково, Николаевка, Новоивановка, Устиновка и Рай-Александровка.
24 июня представитель Народной милиции ЛНР Андрей Марочко заявил «Интерфаксу» о полном контроле над населенными пунктами Золотое и Горское (к востоку от Лисичанска).
Фото: Илья Питалев / РИА Новости
23 июня Андрей Марочко в эфире телеканала «Россия 1» заявил, что силы ЛНР и российские подразделения вошли в населенный пункт Катериновка (к юго-западу от Золотого).
В этот же день посол ЛНР в России Родион Мирошник сообщил о зачистке поселка Лоскутовка (к югу от Лисичанска).
22 июня Народная милиция ЛНР сообщила о занятии поселка Метелкино (рядом с Северодонецком).
20 июня помощник главы МВД ЛНР Виталий Киселев заявил о занятии поселка Тошковка (к юго-востоку от Лисичанска).
10 июня штаб территориальной обороны ДНР сообщил, что Народная милиция при огневой поддержке российской армии установила полный контроль над четырьмя селами: Триполье, Владимировка, Ставки и Липовое.
7 июня Сергей Шойгу сообщил о занятии Святогорска, Ямполя, Дробышево, Яровой, Кировска, а также жилых кварталов Северодонецка.
6 июня штаб территориальной обороны ДНР сообщил об установлении контроля над населенными пунктами Покровское и Зеленый Гай (в районе Угледара).
3 июня в штабе территориальной обороны ДНР сообщили об установлении контроля над населенными пунктами Яровая и Щурово.
30 мая власти ДНР сообщили о продвижении Народной милиции к Славянску и занятии населенных пунктов Диброва и Старый Караван (к западу от Лимана).
28 мая российское Минобороны сообщило, что город Красный Лиман (Лиман) в Донецкой области взят под контроль российскими военными.
24 мая флаг России был поднят над зданием мэрии города Светлодарск (к северо-востоку от Горловки).
Фото: zakharevich_igor / Telegram
21 мая сдались в плен украинские подразделения, оборонявшие металлургический завод «Азовсталь» в Мариуполе.
Фото: Алексей Куденко / РИА Новости
Мариуполь был блокирован российскими войсками и Народной милицией ДНР еще 28 февраля. Спустя месяц украинские подразделения, в том числе полк «Азов» (его символика в России признана экстремистской и запрещена), были блокированы на территории металлургического комбината «Азовсталь». От штурма завода с многочисленными подземными сооружениями было решено отказаться. Попытки украинских военных эвакуировать блокированные подразделения завершились неудачей, после чего начались переговоры о сдаче.
17 мая представитель Народной милиции ЛНР сообщил, что подразделениям республики удалось занять село Орехово (к востоку от города Золотое).
16 мая части ЛНР заняли село Новозвановка (к северу от города Попасная).
12 мая в оборонном ведомстве ЛНР сообщили, что Народная милиция оттеснила украинских военнослужащих от Попасной.
Как пострадала Попасная в ЛНР от боевых действий. Видео
11 мая глава Чечни Рамзан Кадыров сообщил, что в ходе совместного наступления Народной милиции ЛНР и подразделения «Ахмат» заняты населенные пункты Южный и Воеводовка, пороховой завод «Заря», а также несколько дачных поселков к северу и северо-западу от Северодонецка.
5 мая Народная милиция ДНР заняла село Троицкое, из которого велись обстрелы Горловки.
25 апреля Народная милиция ЛНР сообщила о занятии села Новотошковское (к северо-востоку от Попасной).
16 апреля Министерство обороны сообщило о взятии под контроль в Мариуполе Металлургического завода имени Ильича.
11 апреля Минобороны России сообщило о занятии в Харьковской области села Каменка. В военном ведомстве отметили, что это был «один из самых укрепленных рубежей обороны в Изюмском районе».
1 апреля на Украине появились первые публикации об обнаружении тел мирных жителей в городе Буча Киевской области, который ранее покинули российские войска. Затем в Киеве заявили о гибели сотен жителей Бучи и обвинили в случившемся российских военнослужащих. В Москве произошедшее назвали постановкой, указав на целый ряд несоответствий. После событий в Буче переговорный процесс между Россией и Украиной фактически был заморожен.
31 марта Минобороны сообщило о полном занятии населенного пункта Золотая Нива. Российские подразделения форсировали реку Кашлагач.
В этот же день подразделения ЛНР завершили зачистку Житловки.
29 марта российская армия начала покидать Киевскую, Черниговскую и Сумскую области. В Кремле это назвали жестом доброй воли в интересах переговорного процесса.
В течение марта проходили переговоры между делегациями России и Украины. Изначально встречи проводились в Белоруссии, а последняя встреча состоялась в Стамбуле 29 марта. По ее результатам глава российской делегации Владимир Мединский сообщил о согласовании целого ряда позиций, а замминистра обороны Александр Фомин пообещал «кардинальное сокращение военной активности на киевском и черниговском направлениях».
28 марта Народная милиция ЛНР овладела населенными пунктами Ивановка, Новосадовое, выйдя на рубеж Новолюблино — Терны.
25 марта Минобороны сообщило о занятии населенных пунктов Батманка, Михайловка, Красный Партизан, Ставки и Троицкое.
24 марта Минобороны заявило, что российские вооруженные силы взяли под полный контроль город Изюм в Харьковской области.
22 марта Минобороны заявило о потере Киевом контроля над десятью населенными пунктами. Российские военные заняли поселок Урожайное в Донецкой области, силы Донецкой народной республики взяли четыре населенных пункта: Славное, Трудовской, Шахта Челюскинцев и Марьинка. Еще пять украинских населенных пунктов перешли под контроль Народной милиции ЛНР: Калиново, Новоалександровка, Степное и Богуславское.
13 марта Вооруженные силы России установили полный контроль над населенными пунктами Павловка, Никольское, Благодатное, Водяновка, Владимировка.
11 марта Народная милиция ДНР овладела Волновахой.
9 марта в Минобороны России сообщили, что подразделения российских вооруженных сил взяли под контроль населенные пункты Красная Поляна и Степное. Народная милиция ЛНР заняла Грачево, Народная милиция ДНР — Сигнальное, Тарамчук, Еленовку и Трудовое.
4 марта Минобороны России сообщило, что Народная милиция ЛНР заняла несколько населенных пунктов: Шандриголово, Александровку, Дерилово, Среднее, Лозовое. Народная милиция ДНР заняла четыре населенных пункта: Розовку, Труженку, Знаменовку и Пузатое. Российские войска заняли несколько населенных пунктов в Запорожской области, в том числе город Пологи.
3 марта Министерство обороны России сообщило о занятии города Балаклея (Харьковская область). Помимо этого российские войска взяли под контроль несколько населенных пунктов в Запорожской области, среди которых Синяя Гора, Сладкая Балка, Ильченково. Силы ДНР, окружившие Мариуполь, заняли село Водяное, а также Сартану и Виноградное.
Фото: РИА Новости
2 марта Минобороны России сообщило об установлении контроля над Херсоном, а также Васильевкой и Токмаком (Запорожская область). В Луганской области Народная милиция ЛНР установила контроль над городами Старобельск и Сватово, а также райцентром Новоайдар.
27 февраля Минобороны России сообщило о блокировании Херсона и Бердянска, а также о занятии Геническа (Херсонская область). Народная милиция ЛНР заняла населенные пункты Новоахтырка, Смоляниново (к востоку от Северодонецка), Станицу Луганскую.
26 февраля Минобороны России отчиталось, что российские войска за ночь взяли под полный контроль Мелитополь.
25 февраля в Минобороны заявили о занятии острова Змеиный в Одесской области Украины. Гарнизон Змеиного сложил оружие.
24 февраля Погранслужба Украины сообщила об атаках и обстрелах по всей границе от Житомирской области (граничащей только с Белоруссией) до Луганской, а также из Крыма. Ракетным обстрелам подверглись военные объекты по всей территории Украины. Позже стало известно, что российская армия пересекла границу в Киевской, Черниговской, Сумской, Харьковской и Херсонской областях. Одновременно Народная милиция ЛНР начала атаки на города Счастье и Станица Луганская к северу от Луганска, а Народная милиция ДНР начала наступление на Мариуполь и Волноваху.
По требованию Роскомнадзора РБК приводит данные о деталях военной операции на Украине на основании информации российских официальных источников.
Страны НАТО и их союзники с начала спецоперации наращивали поставки оружия Киеву. Если изначально речь шла о касках, бронежилетах, беспилотных летательных аппаратах (турецких Bayraktar), то затем на Украину стали отправлять старое советское вооружение, которое до сих пор хранилось на складах в Польше, Словакии, Чехии.
С течением времени на Украину стали поставлять и вооружение, которое производилось в странах НАТО и Австралии. Речь шла о бронемашинах (британских Saxon, австралийских Bushmaster), противотанковых комплексах (Javelin, NLAW), артиллерийских установках (французских Caesar, американских HIMARS).
Подробно о военных поставках Украине можно прочитать здесь.
Пакеты санкций против России начали вводить еще до начала спецоперации на Украине, после признания ДНР и ЛНР. Об ограничениях объявили США, Канада, ЕС, Япония, Австралия, Новая Зеландия, Великобритания, Швейцария, Исландия и ряд других стран.
Всего было шесть волн санкций. США, Великобритания, ЕС заявляли, что координируют усилия по ослаблению российской экономики, однако не всегда ограничения вводились синхронно. Были и определенные отличия, которые на Западе объясняли нежеланием навредить собственным экономикам. Различия были и в персональных санкциях. Например, США ввели ограничения в отношении 663 человек, а Великобритания — в отношении более 1 тыс. бизнесменов, политиков, военных, сотрудников СМИ, людей, которых в Лондоне посчитали близкими к Путину.
Первая волна касалась банков, финансирующих военную промышленность, депутатов Госдумы, поддержавших признание ДНР и ЛНР. Под санкции также попал ВЭБ, который является кредитором множества компаний, оператором нацпроектов, куратором других институтов развития и управляющий пенсионными накоплениями почти 40 млн россиян.
Вторая волна санкций началась уже после начала спецоперации. Великобритания ввела ограничения в отношении 100 физических лиц и компаний, в том числе ВТБ, «Ростеха» и «Аэрофлота». Были ограничены суммы вкладов и депозитов для граждан России в банках Великобритании до £50 тыс. США ограничили все операции с новыми долговыми обязательствами со сроком погашения более 14 дней целого ряда компаний (в том числе «Газпрома», РЖД, Альфа-банка). Персональные санкции коснулись Владимира Путина, Сергея Лаврова, Сергея Шойгу.
ЕС ввел ограничения в том числе против РЖД, Уралвагонзавода, КАМАЗа, Альфа-банка, «Открытия», банка «Россия» и Промсвязьбанка. Банкам запретили продавать клиентам из России ценные бумаги, номинированные в евро, а также проводить листинг акций любых российских госкомпаний (с долей государства свыше 50%) на европейских биржах. Помимо этого было запрещено страхование российских самолетов, введен запрет на продажу и лизинг лайнеров, запчастей и оборудования российским авиакомпаниям.
К 10 марта воздушное пространство для российских самолетов закрыли ЕС, Великобритания, Босния, Норвегия, США, Швейцария, Черногория, Великобритания. Чуть позже это сделала и Канада. От рейсов в Россию отказался целый ряд авиакомпаний.
Третья санкционная волна началась 26 февраля. Были заморожены резервы Банка России в странах G7, пять банков были отключены от системы SWIFT. Великобритания закрыла порты для российских судов, а также экспорт в Россию высокотехнологичных товаров. На территории ЕС и Великобритании были заморожены активы целого ряда бизнесменов, в том числе Игоря Сечина, Михаила Фридмана, Петра Авена.
2 марта ЕС запретил продавать, поставлять, передавать и экспортировать в Россию банкноты евро.
8 марта США объявили о запрете поставок нефти, газа и угля из России.
C 10 марта работу в России прекратили Visa и Mastercard. Владельцы российских карт этих платежных систем потеряли возможность расплачиваться ими за пределами страны или оплачивать покупки в зарубежных магазинах.
10 марта Великобритания ввела санкции в том числе против Романа Абрамовича, Андрея Костина, Олега Дерипаски. Их активы были заморожены, британским гражданам запретили с ними сотрудничать.
11 марта США запретили поставку в Россию долларовых банкнот, ввоз из России алкоголя, морепродуктов.
12 марта Бермудские Острова аннулировали летные сертификаты 740 зарегистрированных там самолетов российских авиакомпаний.
Четвертая волна санкций началась 15 марта. Был запрещен ввоз в ЕС российской сталелитейной продукции, запрещен экспорт предметов роскоши в Россию.
4 апреля США запретили России использовать замороженные в США резервы для выплат по госдолгу.
5 апреля был объявлен очередной пакет санкций Евросоюза. Так, ЕС запретил импорт российского угля, транзакции с крупнейшими банками. Российским кораблям запретили входить в порты ЕС (исключение сделали для перевозчиков продуктов, гуманитарных и энергетических грузов).
6 апреля США запретили гражданам вести бизнес с Альфа-банком и Сбербанком, а также новые инвестиции в Россию.
9 апреля ЕС ограничил импорт удобрений из России.
11 апреля Комитет по безопасности полетов Евросоюза внес 21 российскую авиакомпанию в список перевозчиков, которым запрещены или для которых ограничены полеты в пределах ЕС, поскольку эти перевозчики не соответствуют международным стандартам безопасности.
12 апреля Великобритания запретила импорт российского чугуна и стали.
3 июня ЕС объявил о шестом пакете санкций. Под ограничения попали в том числе КАМАЗ, «Татнефть», Национальный расчетный депозитарий. От SWIFT были отключены Сбербанк, Россельхозбанк, МКБ. Помимо этого ЕС пообещал в течение шести месяцев отказаться от импорта российской нефти, в течение восьми месяцев — от импорта нефтепродуктов.
Шестой пакет санкций Евросоюз обсуждал больше месяца. Несколько стран указывали, что не готовы пойти на жесткий отказ от российских энергоносителей, поскольку это нанесло бы удар по их экономикам.
16 июня Великобритания ввела санкции в том числе против патриарха Московского и всея Руси Кирилла.
28 июня США ввели запрет на импорт российского золота.
8 июля Канада ввела санкции в отношении патриарха Кирилла. Помимо этого под ограничения попали Роскомнадзор, новостное агентство Regnum, ведущая «Первого канала» Екатерина Андреева.
Video
Президент России Владимир Путин утром 24 февраля выступил с экстренным обращением и объявил о начале специальной военной операции в Донбассе. «Ее [операции] цель — защита людей, которые на протяжении восьми лет подвергаются издевательствам, геноциду со стороны киевского режима», — подчеркнул он.
По его словам, Россия не может чувствовать себя в безопасности и существовать с постоянной угрозой со стороны Украины. Он подчеркнул, что военная операция в Донбассе будет проводиться в соответствии с Уставом ООН, решением Совета Федерации и договорами с ДНР и ЛНР.
США и их союзники хотят отложить пересмотр потолка цен на российскую нефть до марта, сохранив его пока на изначальном уровне $60, сообщает Bloomberg. В то же время Эстония, Литва и Польша, как и прежде, настаивают на снижении лимита, считая его слишком высоким по сравнению с текущими рыночными ценами.
| Формат vk 4D View Pro | Строка | . 4ВП | Формат 4D View Pro (формат по умолчанию) |
| параметры вставки буфера обмена vk все | Лонгинт | 0 | Вставляет все объекты данных, включая значения, форматирование и формулы. |
| параметры вставки буфера обмена vk форматирование | Лонгинт | 2 | Вставляет только форматирование. |
| формулы для вставки буфера обмена vk | Лонгинт | 3 | Вставляет только формулы. |
| параметры вставки в буфер обмена vk формулы и форматирование | Лонгинт | 5 | Вставляет формулы и форматирование. |
| значения параметров вставки из буфера обмена vk | Лонгинт | 1 | Вставляет только значения. |
| параметры вставки буфера обмена vk и форматирование | Лонгинт | 4 | Вставляет значения и форматирование. |
| текущий лист ВК | Лонгинт | -1 | Обозначает текущий лист области 4D View Pro (по умолчанию) |
| vk найти флаг точное совпадение | Лонгинт | 2 | Указывает, что все содержимое ячейки должно полностью соответствовать искомому значению. |
| vk найти флаг игнорировать регистр | Лонгинт | 1 | Указывает, что заглавные и строчные буквы считаются одинаковыми. |
| вк найти флаг нет | Лонгинт | 0 | Указывает, что флаги поиска не учитываются. (по умолчанию) |
| VK найти флаг использовать подстановочные знаки | Лонгинт | 4 | Указывает, что в строке поиска можно использовать подстановочные знаки (*,?). Подстановочные знаки можно использовать в любом сравнении строк для соответствия любому количеству символов.
|
| vk найти порядок по столбцам | Лонгинт | 1 | Указывает, что поиск выполняется по столбцам. |
| вк поиск по строкам | Лонгинт | 0 | Указывает, что поиск выполняется по строкам. Каждый столбец строки просматривается, прежде чем поиск продолжится до следующей строки. (по умолчанию) |
| vk найти целевую формулу | Лонгинт | 8 | Указывает, что поиск выполняется в формуле ячейки. |
| vk найти целевой тег | Лонгинт | 4 | Указывает, что поиск выполняется в теге ячейки. |
| vk найти целевой текст | Лонгинт | 1 | Указывает, что поиск выполняется в тексте ячейки. |
| размер шрифта vk большой | Строка | «большой» | Крупный текст |
| размер шрифта vk больше | Строка | «больше» | Крупный текст |
| средний размер шрифта vk | Строка | «средний» | Текст среднего размера |
| размер шрифта vk маленький | Строка | «маленький» | Мелкий текст |
| vk размер шрифта меньше | Строка | «меньше» | Меньший текст |
| размер шрифта vk x большой | Строка | «большой» | Очень крупный текст |
| размер шрифта vk x маленький | Строка | «х-маленький» | Очень мелкий текст |
| размер шрифта вк xx большой | Строка | «хх-большой» | Очень крупный текст |
| размер шрифта vk xx маленький | Строка | «хх-маленький» | Очень мелкий текст |
| стиль шрифта vk курсив | Строка | «курсив» | Курсив |
| шрифт vk наклонный | Строка | «косой» | Наклонный текст |
| вариант шрифта vk маленькие заглавные | Строка | «маленькие заглавные буквы» | Текст отображается маленькими заглавными буквами. |
| вес шрифта вк 100 | Строка | «100» | Тонкий текст |
| вес шрифта вк 200 | Строка | «200» | Очень светлый текст |
| вес шрифта вк 300 | Строка | «300» | Светлый текст |
| вес шрифта вк 400 | Строка | «400» | Текст нормального веса |
| вес шрифта вк 500 | Строка | «500» | Текст среднего веса |
| вес шрифта вк 600 | Строка | «600» | Полужирный текст |
| вес шрифта вк 700 | Строка | «700» | Жирный текст. |
| вес шрифта вк 800 | Строка | «800» | Очень жирный текст |
| вес шрифта вк 900 | Строка | «900» | Очень жирный текст |
| вес шрифта vk полужирный | Строка | «жирный» | Жирный текст. |
| шрифт vk жирнее | Строка | «смелее» | Более жирный текст |
| шрифт vk легче | Строка | «зажигалка» | Светлый текст |
| автоматический текст заголовка vk пустой | Лонгинт | 0 | Отображает пробелы в заголовках. |
| автотекст писем в заголовке вк | Лонгинт | 2 | Отображает буквы в заголовках. |
| номера автотекста в заголовке vk | Лонгинт | 1 | Отображает числа в заголовках. |
| vk горизонтальное выравнивание по центру | Лонгинт | 1 | Содержимое ячейки выровнено по центру по горизонтали |
| vk горизонтальное выравнивание общее | Лонгинт | 3 | Содержимое ячейки выравнивается по горизонтали в соответствии с типом значения |
| vk горизонтальное выравнивание по левому краю | Лонгинт | 0 | Содержимое ячейки горизонтально выровнено по левому краю |
| vk горизонтальное выравнивание по правому краю | Лонгинт | 2 | Содержимое ячейки горизонтально выровнено по правому краю |
| центр компоновки изображений vk | Лонгинт | 1 | Фоновое изображение отображается в центре 4D View Pro ara |
| макет изображения vk нет | Лонгинт | 3 | Фоновое изображение исходного размера отображается в верхнем левом углу области 4D View Pro |
| vk image layout stretch | Лонгинт | 0 | Фоновое изображение заполняет всю область 4D View Pro |
| vk макет изображения масштаб | Лонгинт | 2 | Фоновое изображение отображается с исходным соотношением сторон |
| выравнивание этикетки vk снизу по центру | Лонгинт | 4 | Этикетка ячейки выровнена по центру снизу |
| выравнивание метки vk внизу слева | Лонгинт | 3 | Метка ячейки выровнена по левому краю внизу |
| выравнивание ярлыка vk внизу справа | Лонгинт | 5 | Этикетка ячейки выровнена по правому краю внизу |
| выравнивание этикетки vk по центру вверху | Лонгинт | 1 | Метка ячейки выровнена по центру сверху |
| выравнивание метки vk вверху слева | Лонгинт | 0 | Метка ячейки выровнена по левому краю вверху |
| выравнивание метки vk справа вверху | Лонгинт | 2 | Метка ячейки выровнена по правому краю |
| видимость ярлыка вк авто | Лонгинт | 2 | Отображает водяной знак в области заполнения (если в ячейке есть значение) или в ячейке (если в ячейке нет значения) |
| видимость ярлыка вк скрыта | Лонгинт | 1 | Отображает водяной знак в области ячейки на основе условия значения |
| видимость ярлыка vk | Лонгинт | 0 | Водяной знак всегда отображается в области заполнения, независимо от значения ячейки |
| пунктирная линия в стиле vk | Лонгинт | 9 | Линия, состоящая из одного тире и одной точки |
| стиль линии vk тире точка точка | Лонгинт | 11 | Линия, состоящая из одного тире и двух точек |
| пунктирная линия vk | Лонгинт | 3 | Линия из штрихов |
| пунктирная линия vk | Лонгинт | 4 | Линия из точек |
| vk line стиль двойной | Лонгинт | 6 | Линия из двух линий |
| стиль линии vk пустой | Лонгинт | 0 | Простая линия без стиля |
| волосы в стиле vk line | Лонгинт | 7 | Линия из точек. |
| vk line style средний | Лонгинт | 2 | Прочная леска средней толщины |
| VK линия, средний штрих, пунктирная точка | Лонгинт | 10 | Линия средней толщины, состоящая из одного тире и одной точки |
| стиль линии vk средний тире многоточие | Лонгинт | 12 | Линия средней толщины, состоящая из одного тире и двух точек |
| стиль линии vk средний пунктир | Лонгинт | 8 | Линия средней толщины из штрихов |
| стиль линии vk наклонный штрихпунктирная точка | Лонгинт | 13 | Линия, состоящая из наклонной черты и наклонной точки |
| толстая линия vk | Лонгинт | 5 | Толстая сплошная линия |
| тонкая линия vk | Лонгинт | 1 | Легкая сплошная линия |
| vk Формат MS Excel | Строка | . xlsx | Формат Microsoft Excel |
| vk шаблон полная дата время | Строка | «_fullDateTimePattern_» | Формат ISO 8601 для полной даты и времени в текущей локализации. Шаблон по умолчанию для США: «dddd, dd MMMM yyyy HH:mm:ss» |
| vk шаблон с длинной датой | Строка | «_longDatePattern_» | Формат ISO 8601 для полной даты в текущей локализации. Шаблон по умолчанию для США: «dddd, dd MMMM yyyy» |
| шаблон вк давно | Строка | «_longTimePattern_» | Формат ISO 8601 для времени в текущей локализации. Шаблон по умолчанию для США: «ЧЧ:мм:сс» |
| шаблон вк месяц день | Строка | «_monthDayPattern_» | Формат ISO 8601 для месяца и дня в текущей локализации. Шаблон по умолчанию для США: «ММММ дд» |
| vk шаблон короткая дата | Строка | «_shortDatePattern_» | Сокращенный формат ISO 8601 для даты в текущей локализации. Шаблон по умолчанию для США: «ММ/дд/гггг» |
| шаблон вк короткое время | Строка | «_shortTimePattern_» | Сокращенный формат ISO 8601 для времени в текущей локализации. Шаблон по умолчанию для США: «ЧЧ:мм» |
| vk шаблон сортировки по дате и времени | Строка | «_sortableDateTimePattern_» | Формат ISO 8601 для даты и времени в текущей локализации, который можно сортировать. Шаблон по умолчанию для США: «гггг\’-\’ММ\’-\’дд\’Т\’ЧЧ\’:\’мм\’:\’сс» |
| vk шаблон универсальная сортируемая дата и время | Строка | «_universalSortableDateTimePattern_» | Формат ISO 8601 для даты и времени в текущей локализации с использованием UTC, который можно сортировать. Шаблон по умолчанию для США: «yyyy\’-\’MM\’-\’dd HH\’:\’mm\’:\’ss\’Z\'» |
| vk шаблон год месяц | Строка | «_yearMonthPattern_» | Формат ISO 8601 для месяца и года в текущей локализации. Шаблон по умолчанию для США: «yyyy MMMM» |
| вк pdf формат | Строка | Формат PDF | |
| vk позиция нижняя | Лонгинт | 2 | Вертикальное выравнивание по низу ячейки или строки |
| вк позиция центр | Лонгинт | 1 | Выравнивание по центру. Выравнивание будет осуществляться по границе ячейки, строки или столбца в соответствии с указанным положением просмотра:
|
| вк позиция слева | Лонгинт | 0 | Выравнивание по горизонтали слева от ячейки или столбца |
| vk позиция ближайшая | Лонгинт | 3 | Выравнивание по ближайшему пределу (сверху, снизу, слева, справа, по центру).
|
| вк позиция справа | Лонгинт | 2 | Выравнивание по горизонтали справа от ячейки или столбца |
| вк позиция сверху | Лонгинт | 0 | Вертикальное выравнивание по верху ячейки или строки |
| vk print по центру оба | Лонгинт | 3 | Печать по центру страницы по горизонтали и вертикали |
| vk print центрирование по горизонтали | Лонгинт | 1 | Печать выполняется по центру страницы по горизонтали. |
| vk print центрирование нет | Лонгинт | 0 | Печать не по центру. (по умолчанию) |
| vk print центрирование по вертикали | Лонгинт | 2 | Печать выполняется по центру страницы по вертикали. |
| vk print page order auto | Лонгинт | 0 | Порядок печати определяется автоматически. (по умолчанию) |
| порядок страниц печати vk вниз, затем выше | Лонгинт | 1 | Страницы печатаются в порядке убывания, затем поперек. |
| vk порядок страниц печати сверху вниз | Лонгинт | 2 | Страницы печатаются поперек, затем в порядке убывания. |
| ориентация страницы vk для печати альбомная | Лонгинт | 2 | Альбомная ориентация |
| ориентация страницы vk для печати книжная | Лонгинт | 1 | Портретная ориентация. (по умолчанию) |
| вк печать видимость скрыть | Лонгинт | 1 | Заголовок не виден. |
| vk print видимость наследовать | Лонгинт | 0 | Наследует настройки листа (по умолчанию) |
| вк печать видимость показать | Лонгинт | 2 | Заголовок виден на каждой странице. |
| вк печать видимость показать один раз | Лонгинт | 3 | Заголовок виден один раз. |
| оформление текста vk двойное подчеркивание | Лонгинт | 8 | Отображает двойную строку под текстом |
| оформление текста vk через строку | Лонгинт | 2 | Отображает строку через текст |
| украшение текста вк нет | Лонгинт | 0 | Отображает текст без оформления |
| vk украшение текста над чертой | Лонгинт | 4 | Отображает строку над текстом |
| оформление текста ВК подчеркивание | Лонгинт | 1 | Отображает одну строку под текстом |
| вертикальное выравнивание по нижнему краю | Лонгинт | 2 | Содержимое ячейки выравнивается вертикально по нижнему краю |
| vk вертикальное выравнивание по центру | Лонгинт | 1 | Содержимое ячейки вертикально выровнено по центру |
| vk вертикальное выравнивание вверху | Лонгинт | 0 | Содержимое ячейки выровнено вертикально вверх |
| рабочая тетрадь вк | Лонгинт | -2 | Обозначает всю рабочую книгу области 4D View Pro. |
CvkR представляет собой репрессор транскрипции MerR-типа систем транспозаз, связанных с CRISPR, типа V-K класса 2
Введение
Нативные кластеризованные регулярно расположенные короткие палиндромные повторы (CRISPR) и CRISPR-ассоциированные (Cas) белки хорошо охарактеризованы в отношении их функции как основанные на РНК адаптивные и наследуемые иммунные системы, присутствующие у многих бактерий и архей 1,2,3,4,5,6 . Несколько генетических подходов, разработанных на основе этих нативных систем CRISPR-Cas, стали популярными для манипулирования экспрессией генов и редактирования генома 7,8,9 .
Системы CRISPR-Cas очень разнообразны и подразделяются на 2 класса, 6 типов и 33 подтипа 10 . Недавно была обнаружена замечательная группа производных, которые представляют собой гибриды Tn7-подобных транспозонов и систем CRISPR, кодирующих эффекторы Cas12k с естественными инактивированными нуклеазными доменами 11 или кодирующие каскадные комплексы без компонента нуклеазы Cas3 12,13 . Соответствующие системы CRISPR, связанные с транспозонами, включают системы класса 1 типа I-F, I-B и класса 2 типа V-K 13,14 . Эти системы, называемые CRISPR-ассоциированными транспозонами (CAST), способны катализировать транспозицию мобильных генетических элементов, управляемых crRNAs, в то время как системы, ассоциированные с типом I-B, используют специальный механизм самонаведения на основе белков TniQ/TnsD 13 .
Система, охарактеризованная на примере Vibrio cholerae , состоит из генов, кодирующих белки CRISPR-Cas подтипа I-F Cas6, Cas7, слитый белок Cas5-Cas8, и генов, кодирующих белки-транспозоны TnsA, TnsB, TnsC и TniQ 12,14 . Сообщалось об одном экземпляре систем CAST типа I-B класса 1 и нескольких типах V-K класса 2 в нескольких различных цианобактериях, которые представляют собой богатый природный ресурс для этих систем 11,13,15 .
Системы V-K CAST, впервые охарактеризованные в Scytonema hofmanni 11 содержат гены, кодирующие субъединицу эффекторного комплекса Cas12k и субъединицы Tn7-подобной транспозазы TnsB, TnsC и TniQ, в то время как tnsA отсутствует. Неожиданным открытием было то, что рибосомный белок S15 рекрутируется в связанный с ДНК-мишенью комплекс Cas12k-transposon 16 . Нацеливание транспозиции с помощью этих систем CAST зависит от взаимодействия ДНК-crРНК, облегчаемого эффекторным белком Cas12k 11 . Затем транспозон TnsC образует спиральные полимеры вокруг ДНК, поддерживаемые связыванием АТФ 17 . Рост в направлении от 5′ к 3′ останавливается за счет связывания TniQ на конце филамента, одновременно соединяющего филамент TnsC с Cas12k. Затем на другом конце филамента Mu-подобная транспозаза TnsB начинает интегрировать транспозон 17 . В дополнение к генам, кодирующим транспозазы и эффекторные белки, все эти системы содержат различное количество генов-карго. Новые генетические подходы были разработаны на основе различных гибридных систем Tn7-CRISPR-Cas 18,19,20 , подчеркнув, что более точное описание таких систем представляет как фундаментальный, так и прикладной интерес.
В то время как парадигма состоит в том, что нативные системы CRISPR-Cas в первую очередь защищают целостность генома от мобильных генетических элементов, системы CAST, похоже, нарушают эту парадигму, поскольку они по определению представляют собой мобильные элементы. Из-за этих противоречивых функций можно ожидать жесткого регулирования этих систем. Действительно, сообщалось, что системы CAST также содержат ген, кодирующий предполагаемый регулятор транскрипции MerR-типа 9.1378 15 . Классические регуляторы MerR-типа уникальным образом модулируют генную экспрессию путем связывания центральной области промотора с нетипичными длинными спейсерами между -35 и -10 элементами 21,22,23 . Однако ассоциация регуляторов MerR с системами CAST не была систематически исследована, и до сих пор ее функция не изучалась экспериментально.
Мы изучили системы CRISPR-Cas у Anabaena ( Nostoc ) sp. PCC 7120 (отсюда: Anabaena 7120), многоклеточная азотфиксирующая модельная цианобактерия с богатой CRISPR хромосомой из одиннадцати CRISPR-подобных повторных спейсерных кассет. Все они расшифрованы 15 , и на основе особенностей родственных эндонуклеаз созревания Cas6 пять из этих массивов были отнесены к типу III-D, а еще пять — к системе CRISPR-Cas типа I-D 24 , в то время как оставшийся массив (CR_9) относится к отдельному типу CRISPR со всеми признаками CAST-системы 15,24 .
Здесь мы сначала тщательно изучаем связь между предполагаемыми регуляторами транскрипции и CAST-системами цианобактерий и обнаруживаем, что они принадлежат к трем разным типам, причем большинство из них представляют собой MerR-подобные регуляторы. Затем мы исследуем MerR-подобный регулятор транскрипции Alr3614, принадлежащий системе Anabaena 7120 CRISPR-ассоциированной транспозазы (AnCAST). Мы установили, что как cas12k ген all3613 , так и merR -подобный ген alr3614 транслируются с мРНК без лидера, и что Alr3614 функционирует как репрессор системы AnCAST. Мы идентифицировали консервативный мотив связывания Alr3614 длиной 15 нуклеотидов с помощью анализов следа ДНКазы I и сдвига в геле, а также биоинформатического анализа. Анализ кристаллической структуры показывает, что Alr3614 принимает уникальный паттерн димеризации и режим упаковки в С-концевой сенсорной области, в то время как ему не хватает узнаваемого сайта связывания эффектора по сравнению с другими членами семейства MerR. Таким образом, Alr3614 является необычным регулятором семейства MerR со структурной точки зрения. Мы предлагаем называть Alr3614 и его гомологи репрессором Cas V-K (CvkR), кодируемым геном cvkR .
Результаты
Архитектура CAST-систем цианобактерий
Исходя из известных компонентов CAST, мы искали консервативные гены и генетические элементы в их окрестностях. Эти элементы включали левый и правый концы транспозона, соседние тРНК, массивы CRISPR и tracrРНК, гены транспозазы ( tnsB, tnsC и tniQ ) и гены в обратной ориентации рядом со стартовым кодоном cas12k . Предполагается, что он кодирует небольшие ДНК-связывающие белки. Мы идентифицировали 118 систем CAST с четкими 9 баллами. 1396 гена cas12k в 88 различных штаммах. Большинство из них было обнаружено у Nostocales (60%), Chroococcales (15%) и Pseudanabaenales (11%), дополненных небольшим количеством систем CAST у цианобактерий Oscillatoriales, Pleurocapsales, Spirulinales и Synechococcales (дополнительные данные 1). . Три дополнительные системы CAST были идентифицированы у неклассифицированных нитчатых цианобактерий (CCT1, CCP2 и 4).
На основе этого анализа мы определили общую структуру системы CAST этого типа (рис. 1) в соответствии с предыдущими анализами 10,11,15 и добавил дополнительную информацию. Левый конец обычно лежит ниже гена тРНК, который направлен к транспозону 12,13,15 . Массив CRISPR всегда следует на небольшом расстоянии в обратной ориентации, в направлении левого конца 11,13 .
Рис. 1: Основное расположение генов в CAST-системах цианобактерий. Транспозон CAST отображается с левого конца (LE) на правый конец (RE). Гены окрашены в соответствии с функцией (зеленый: cas12k , оранжевый: гены транспозазы, голубой: ген регулятора CAST ( cvkR в Anabaena 7120), желтый: участки, из которых транскрибируется некодирующая РНК, темно-серый: гены-карго. Сверху увеличена область от гена тРНК до tracrРНК. Массив CRISPR изображен с повторами (серый) и спейсерами (желтый), отделенными от 17-нуклеотидного якорного спейсера (синий) ниже массива рядом с усеченной последовательностью повторов (R *; ~ 12 нт). Схема выполнена не в масштабе, но указаны расстояния, представляющие особый интерес или упомянутые в тексте.
Изображение в полный размер
Повторы CRISPR в этих системах CAST наиболее консервативны в своих 3′-сегментах 13 и 37 нуклеотидов в длину. За массивом CRISPR следует ген tracrRNA (рис. 1) на той же цепи 11 ,25,26,27 . Основываясь на предыдущем полногеномном картировании сайтов начала транскрипции (TSS) 28 , мы обнаружили tracrRNA TSS 35 нуклеотидов ниже стоп-кодона эффекторного гена CRISPR-Cas cas12k , транскрибируемого в том же направлении (дополнительная рис. 1а). Промотор tracrRNA, по-видимому, консервативен среди различных систем CAST (дополнительная рис. 1b).
Рядом с левым концом усеченный одиночный повтор лежит внутри транспозона ниже по течению, но обычно четко отделен от массива CRISPR 13 (обозначен R * на рис. 1). Непосредственно ниже этого усеченного повтора R* может быть идентифицирована укороченная спейсерная последовательность, обычно состоящая из 17 нуклеотидов. Этот укороченный спейсер соответствует последовательности протоспейсера, расположенной сразу за пределами транспозона рядом с левым концом (рис. 1), обычно внутри гена тРНК 13 . Усеченные последовательности с одним повтором-спейсером, прочитанные ближе к левому концу, показывают консервативный восходящий GTN-PAM, соответствующий предсказанному Cas12k PAM 9.1378 11 . Расстояние от этого PAM до левого конца варьируется от 46 до 82 н., за одним исключением 147 н. (рис. 1). Из-за его консервативного положения в этом сайте и экспериментальных доказательств интеграции CAST посредством распознавания протоспейсеров 13 этот мотив, вероятно, необходим для вставки CAST. Поэтому мы предлагаем термины якорный протоспейсер и якорный спейсер для этих последовательностей (рис. 1).
Глядя с другой стороны транспозона, первые гены рядом с правым концом — это три гена, кодирующие субъединицы транспозазы TnsB, TnsC и TniQ, всегда в таком порядке 11,13,29 , отвернуты от правого конца (рис. 1). Большинство грузовых генов, расположенных между cas12k и tniQ , менее консервативны и различаются по последовательности и функциям. Однако есть одно исключение: ген, который, по прогнозам, кодирует небольшой ДНК-связывающий белок рядом со стартовым кодоном cas12k в обратной ориентации (рис. 1).
Три типа регуляторов связаны с CAST-системами цианобактерий
После идентификации 118 CAST-систем и их соответствующих левого и правого концов (дополнительные данные 1) мы разработали базу данных всех белков, возможно, кодируемых в CAST-транспозонах с минимальная длина 50 аминокислот и наличие трех стартовых кодонов ATG, TTG и GTG. Мы искали эту базу данных по blastP 30 для гомологов Anabaena 7120 MerR-подобного белка Alr3614 (в дальнейшем называемого CvkR), ранее предложенного в качестве возможного регулятора 15 , допускающего максимальное значение e 1e-20. Мы идентифицировали 53 merR -подобных генов, все они расположены выше cas12k в обратной ориентации, аналогично расположению cvkR в Anabaena 7120.
Параллельно мы использовали веб-инструмент NCBI для поиска в базе данных консервативных доменов. 31 для присвоения доменов белкам в нашей базе данных. Изучая белки, кодируемые выше cas12k в обратной ориентации, мы обнаружили десять генов с доменом cl10310 (суперсемейство PHA01623) с неизвестной функцией (в дальнейшем называемых «Arc_1») и шесть генов с доменом cl06769 (RHH_5, лента). -helix-helix; ниже называется «Arc_2»). Дальнейший анализ с помощью HHpred 32 предсказал, что оба набора генов кодируют малые Arc-подобные репрессоры из 53–72 аминокислот, которые относятся к суперсемейству ДНК-связывающих белков лента-спираль-спираль 9. 1378 33,34 . Чтобы идентифицировать дополнительные варианты, мы провели поиск гомологов этих репрессоров Arc в системах CAST (максимальное значение e 1e-20) и идентифицировали в общей сложности 22 гена Arc_1 (выравнивание в дополнительных данных 2) и 11 генов Arc_2 (выравнивание в дополнительных данных 3). ), которые все были расположены перед cas12k в обратной ориентации. Мы обнаружили еще один ген Arc_1, который на расстоянии 1,3 т.п.н. расположен дальше от cas12k , и еще один, лишенный ассоциации с cas12k .
Мы дополнительно исследовали это значение путем поиска дополнительных гомологов этих регуляторов за пределами известных систем CAST в NCBI и идентифицировали 210 уникальных генов (157 CvkR-подобных, 32 Arc_1-подобных и 21 Arc_2-подобных; максимальное значение e 1e -20). Мы искали cas12k в непосредственной близости от этих генов и в 169 случаях обнаружили гена cas12k или его вырожденную версию (дополнительная таблица 1). Мы также искали другие компоненты CAST рядом с регуляторами, что дало 130 левых концевых элементов, 90 массивов CRISPR, 139 генов тРНК, а также 119 локусов tracrRNA, идентифицированных с помощью инструмента CRISPRtracrRNA 27 . Всего в 18 случаях мы не смогли найти компоненты CAST рядом с регулятором. Эта высокая ассоциация с системой CAST подтверждает важность этих регуляторов для транспозона и cas12k .
Таким образом, были идентифицированы три основных типа возможных регуляторов: MerR-подобный белок, названный здесь CvkR, и два различных репрессора Arc, названных здесь Arc_1 и Arc_2.
Чтобы исследовать связь этих различных типов регуляторов с системами CAST в различных таксонах, мы провели филогенетический анализ белков Cas12k и нанесли соответствующие регуляторы на карту полученного филогенетического дерева. Чтобы избежать ошибок, мы исключили, вероятно, усеченные и псевдогены, получив 106 генов (рис. 2, дополнительный рисунок 2, выравнивание в дополнительных данных 4). Этот анализ показал, что три основных типа регуляторов стали ассоциироваться с тремя отдельными группами белков Cas12k, хорошо отделенными друг от друга. Этот результат также свидетельствует об отдельных отдельных событиях, в которых наследственные системы CAST включали соответствующий регулятор. Напротив, отношения между конкретным Cas12k и конкретным таксоном не соответствуют различным порядкам цианобактерий, вероятно, из-за горизонтальной передачи систем CAST.
Идентифицированные 106 полных белков Cas12k были сопоставлены с использованием M-coffee 74,75 и проанализированы с помощью BEAST 76 . Двенадцать случаев вырожденных белков Cas12k были намеренно исключены, чтобы избежать неправильного толкования. Результирующее дерево изображается с ветвями, помеченными их соответствующей апостериорной вероятностью, если ≥0,7. Для лучшего распознавания белки были помечены соответствующим организмом-хозяином (дополнительные сведения см. в дополнительных данных 1). Соответствующий тип регулятора (AR) и таксономический порядок (O) соответствующего организма-хозяина имеют цветовую кодировку. Anabaena 7120 Cas12k (All3613) (NCBI: BAB75312.1) отмечен зеленой пунктирной рамкой. Четыре узла, все ветви которых представляли генов cas12k с аналогичными ассоциированными регуляторными генами и организмами-хозяевами с аналогичным таксономическим порядком, были свернуты и помечены соответствующим количеством генов cas12k . Расширенное филогенетическое дерево изображено на дополнительном рисунке 2, а множественное выравнивание последовательностей, служащее исходными данными для филогенетического анализа, доступно в дополнительных данных 4.
Изображение в полный размер
Длина белков семейства CvkR составляет 139–185 аминокислот, и на основе филогенетического анализа можно выделить две отдельные подгруппы (дополнительный рисунок 3, выравнивание в дополнительных данных 5). Четыре белка CvkR различались еще тем, что представляли слитые белки с доменом рестрикции hsdR из системы ДНК-рестрикции-метилирования (RM) (дополнительный рисунок 3). Системы RM часто встречаются среди генов грузов CAST, но нет никаких доказательств того, что системы RM и CAST работают вместе на механистическом уровне.
Экспрессия CvkR из безлидерной мРНК
CvkR, кодируемый alr3614 , был аннотирован как белок MerR длиной 168 аминокислот. Однако сравнение последовательностей Alr3614 с другими CvkR MerR-типа показало, что аннотация NCBI для этого белка (названного здесь CvkR-L) слишком длинная (поддерживается 49 из 53 гомологов), и фактический белок будет состоять всего из 150 остатков. (здесь называется CvkR-S; рис. 3a). Более того, стартовый кодон для трансляции 150 остатков CvkR-S совпадает с ранее картированным TSS его мРНК 28 , что убедительно свидетельствует о трансляции CvkR-S с мРНК без лидера.
Рис. 3: Безлидерная экспрессия генов cvkR . a Выведенное выравнивание аминокислотных последовательностей 53 гомологов CvkR семейства MerR. Последовательности гомологов CvkR семейства MerR для множественного выравнивания последовательностей были восстановлены из общедоступных баз данных и доступны в дополнительных данных 1. Последовательности были выровнены с использованием MAFFT 87 и визуализированы с помощью UGENE 88 . b Данные дифференциального секвенирования РНК 28 показывают, что TSS cvkR и cas12k располагались внутри кодирующих последовательностей, аннотированных NCBI, при восполнении азота (темно-зеленый) и 8-часовом истощении азота (светло-зеленый). Желтые прямоугольники обозначают соответствующие разделы, не использованные в переводе. Данные показаны как первичное покрытие чтения, сопоставленное с соответствующей областью генома. Каждый TSS представлен стрелкой. c Расстояния между cas12k эффекторных белков и генов-регуляторов нанесены на график в соответствии с соответствующим типом регулятора, CvkR ( n = 51 исследованных независимых систем CAST), Arc_1 ( n = 21) и Arc_2 ( n = 11). Блочные диаграммы окрашены в соответствии с соответствующим ДНК-связывающим доменом (HTH: синий; RHH: красный) и структурированы как центр: медиана; границы поля: верхний и нижний квартили; усы: 1,5× межквартильный размах; точки: отдельные образцы. d Анализы qRT–PCR подтверждают, что cvkR транскрибируется в штаммы Δ cvkR Com. Количества транскриптов cvkR нормализовали по количеству rnpB в качестве внутреннего стандарта. Данные представлены в виде средних значений ± SD ( n = 3 биологически независимых эксперимента). e Вестерн-блот-анализ различных комплементарных штаммов Δ cvkR показывает экспрессию белков CvkR разного размера. Верхняя панель, вестерн-блот против С-концевой метки 3xFLAG; Нижняя панель, окрашивание понсо S показывает, что были загружены равные количества белка (100 мкг). Расчетные молекулярные массы CvkR-S и CvkR-L составили 20,03 кДа и 22,41 кДа соответственно. Было проведено два независимых эксперимента, которые показали согласующиеся результаты. Δ 9Штаммы 1396 cvkR Com-1, 2 и 3 подробно описаны на дополнительном рисунке 4c. Исходные данные предоставляются в виде файла исходных данных.
Изображение в полный размер
Аналогичным образом мы заметили, что TSS cas12k совпадает с первым нуклеотидом предполагаемого стартового кодона для рамки считывания, что дает белок из 639 аминокислот (верно в записи GenBank MBD2276705) вместо белок из 648 остатков (аннотированный в записи GenBank BAB75312. 1). Следовательно, и CvkR, и Cas12k, вероятно, транслируются с безлидерной мРНК (рис. 3b). Межгенный спейсер между двумя скорректированными стартовыми кодонами и соответствующими им TSS имеет длину 82 нуклеотида в Anabaena 7120. Обычно более короткое расстояние между этими двумя родственными генами, чем с другими ассоциированными регуляторами, также наблюдалось для других систем (рис. 3c).
Чтобы проверить беслидерную экспрессию CvkR-S, мы сконструировали делеционный мутант (Δ cvkR ) с использованием инструмента редактирования генома CRISPR-Cas12a (Cpf1) (дополнительный рисунок 4a, b) и три версии комплементационных мутантов (Δ cvkR Com-1, -2 и -3). Для комплементации использовались челночные векторы, которые несли генов cvkR , управляемых индуцируемым медью промотором petE для длинной формы CvkR-L (Δ cvkR Com-1) или короткой формы CvkR-S, либо содержащей лидерную последовательность (Δ cvkR Com- 3) или нет (Δ cvkR Com-2). Для обнаружения метку, кодирующую эпитоп 3xFLAG, сливали со всеми тремя рамками считывания cvkR . Комплементарные плазмиды были введены в фон делеции Δ cvkR и проверены (дополнительная рис. 4c, d). гена cvkR транскрибировали во всех трех конструкциях (рис. 3d). Трансляция кодируемых белков была обнаружена с помощью вестерн-блоттинга с четкими различиями в размерах между CvkR-L и CvkR-S (рис. 3e). Интересно, что в Δ cvkR Com-1 была обнаружена не только форма CvkR-L, но также небольшое количество CvkR-S, что указывает на склонность к инициации трансляции в кодоне 18, даже если мРНК была 5′-удлиненной. Поскольку не было 5’UTR в Δ cvkR Com-1 после P petE , кажется, что стартовый кодон CvkR-S может также служить стартовым сайтом трансляции в сочетании с транскрипцией с сильного промотора. Несоответствие между мРНК cvkR и уровнями белка в Δ cvkR Com-3 означает, что 5’UTR P petE может влиять на скорость транскрипции, стабильность мРНК и/или эффективность трансляции cvkR-S. мРНК. В совокупности мы предоставляем убедительные доказательства того, что CvkR может экспрессироваться из безлидерной мРНК (Δ cvkR Com-2) или стартовый кодон, соответствующий кодону 19 исходной аннотации (Δ cvkR Com-3). Поскольку правильная индуцируемость от промотора P petE наблюдалась только у Δ cvkR Com-2, мы использовали этот штамм для всех последующих экспериментов по комплементации, называя его в дальнейшем Δ cvkR Com.
CvkR влияет на экспрессию ключевых компонентов системы CAST in vivo
Система AnCAST содержит все известные ассоциированные гены и простирается примерно на 21 т.п.н. (рис. 4a). В дополнение к своим основным компонентам CAST он кодирует множество других транспозаз, возможных регуляторов, пару токсин/антитоксин и систему RM. 9Ген 1396 cas12k ( all3613 ) лежит вверх по течению, обратно к cvkR . Чтобы проанализировать функцию CvkR in vivo, отсутствие или сверхэкспрессия CvkR в Δ cvkR или Δ cvkR Com были сначала подтверждены с помощью нозерн-гибридизации (дополнительная рис. 5a) и вестерн-блоттинга (дополнительная рис. 5b) соответственно после индукция транскрипции cvkR в Δ cvkR Com из промотора petE , индуцируемого медью. Затем проводят нозерн-гибридизацию тотальной РНК из тройных клонов WT, Δ cvkR и Δ cvkR Com с одноцепочечными РНК-зондами. Специфические сигналы для мРНК cvkR были обнаружены в Δ cvkR Com (рис. 4b). Нозерн-гибридизация против tracrRNA и массива AnCAST CRISPR давала сигналы <500 nt, которые были сильно увеличены в Δ cvkR по сравнению с контролем дикого типа и уменьшились по интенсивности в комплементарном штамме Δ cvkR Com (рис. 4c, d). ). Мы также обнаружили повышенную интенсивность сигнала для 9мРНК 1396 cas12k у Δ cvkR (рис. 4e), значительно выше, чем у WT и Δ cvkR Com (рис. 4f). Эти данные предоставляют прямые доказательства того, что CvkR прямо или косвенно регулирует количество транскриптов, происходящих от промотора tracrRNA, и мРНК cas12k .
a Гены в пределах 24,9AnCAST из 63 нуклеотидов включает cvkR , кодирующий регулятор транскрипции, cas12k , кодирующий эффектор, несколько генов-карго и Tn7. Серые треугольники обозначают гибридизационные зонды, использованные в b – e . Гены RM окрашены в пурпурный цвет, модуль токсин-антитоксин — в черный цвет, другие функции генов показаны на рис. 1. b — e Северные гибридизации для cvkR мРНК, массива CRISPR, tracrRNA и cas12k , как указано. Треугольники указывают ожидаемую длину для cvkR , для массива CRISPR (полная длина ~ 250 нуклеотидов, две единицы повтора-спейсера ~ 140 нуклеотидов, частично процессированные две единицы повтора-спейсера ~ 100 нуклеотидов, один повтор-спейсер 70 нуклеотидов), tracrRNA и cas12k , ген размером 1,92 т. п.н. Гели и мембраны проверяли на равную загрузку окрашиванием бромистым этидием и гибридизацией с 120 нуклеотидной 5S рРНК (в b и d одна и та же мембрана была гибридизирована). Лестницу одноцепочечной РНК нижнего диапазона (NEB) использовали в качестве маркера размера в b и 9.1602 d , лестницы РНК низкого и высокого диапазона RiboRuler (Thermo Fisher Scientific) в c и e . Двадцать микрограмм тотальной РНК WT или соответствующих мутантов разделяли на 10% ПАА-мочевина ( b – d ) или 1,5% формальдегид-агарозный гель ( e ). f Интенсивность сигналов мРНК cas12k , нормализованных к 5S рРНК. Данные представлены в виде средних значений ± SD ( n = 3 биологически независимых образца). Двусторонний t-критерий равной дисперсии показал различные уровни экспрессии для каждого штамма. г График вулкана Δ cvkR и Δ cvkR Анализ транскриптома Com. Горизонтальная линия отмечает пороговое значение p , равное 0,01, вертикальные линии — пороговое значение кратности изменения │log 2 │ ≥ 1. Транскрипты выше этих порогов показали значительное снижение (синий) или повышение (красный) Δ cvkR . (подробности в дополнительной таблице 3). h Наиболее дифференциально экспрессируемая область при анализе микрочипов принадлежит системе AnCAST. Отдельные датчики обозначены горизонтальными полосами, окрашенными в голубой цвет для Δ cvkR и темно-синим цветом для Δ cvkR Com. Анализы, показанные в b — e , были выполнены дважды с согласованными результатами. Исходные данные предоставляются в виде файла исходных данных.
Изображение полного размера
Более того, наличие транскриптов >400 нт указывало на то, что tracrRNA и массив CRISPR транскрибируются в более длинный предшественник, который впоследствии процессируется в основные накапливающиеся фрагменты длиной ~150 и ~210 нт соответственно. Эти длины фрагментов согласуются с предыдущими результатами дифференциальной РНК-seq 9.1378 28 и обширные данные транскриптома 24 , которые предполагают, что массив AnCAST CRISPR не будет иметь своего собственного специфического промотора. Вместо этого объединенный предшественник массива tracrRNA-CRISPR транскрибируется с TSS, расположенной на 35 нуклеотидов ниже кодирующей последовательности cas12k , в положении 4362990 на обратной цепи, на 413 нуклеотидов выше первого повтора массива CRISPR 15 (Дополнительный рис. 1а). Тем не менее, следует отметить, что массив CRISPR показал гораздо более обильные сигналы в области Δ 9Делеционный мутант 1396 cvkR по сравнению с WT и Δ cvkR Com, в то время как tracrРНК также хорошо выявлялась у WT и Δ cvkR Com (рис. 4c, d). Таким образом, существуют дополнительные факторы, вероятно действующие на процессинг и стабилизацию транскриптов, полученных из массива.
Чтобы исследовать глобальные транскриптомные изменения после делеции cvkR , был проведен анализ микрочипов с использованием проверенных выше штаммов Δ cvkR и Δ cvkR Com с делецией и комплементацией (дополнительная рис. 4). Используемые микроматрицы охватывают все гены, кодирующие белок, а также некодирующие РНК, и были специально разработаны для прямой гибридизации меченой тотальной РНК без предварительного преобразования в кДНК 9. 1378 35 . Следовательно, они позволяют напрямую определять уровни транскриптов tracrRNA и массива CRISPR. Микроматричный анализ выявил небольшое количество генов с нарушенной регуляцией. Тринадцать признаков были значительно усилены (семь изменений в генах, кодирующих белок, и шесть других транскриптов), а 8 были значительно подавлены (шесть генов, кодирующих белок, и два других транскрипта) в Δ cvkR по сравнению с Δ cvkR Com (рис. 4g). ). В дополнение к генам AnCAST cas12k и tnsB ( all3630 ), tracrRNA и массив CRISPR активировались (рис. 4h) после делеции cvkR . Следовательно, роль CvkR как регулятора системы AnCAST была подтверждена и расширена.
Дифференциально экспрессируемые гены за пределами системы AnCAST включали L-матрицу, кластер скрытых генов тРНК, имеющий значение для выживания в условиях трансляционного стресса 36 , который был усилен здесь в Δ cvkR Com (рис. 4g), обратный регуляции наблюдается в течение cas12k и tnsB . Другими дифференциально регулируемыми генами были соседний ген L-массива all8564 , кодирующий самонаводящуюся эндонуклеазу типа HNH, rtcB , кодирующий РНК-лигазу, связанную с репарацией РНК 36 , а также alr0739 и alr0739 и alr0740
Набор данных всех транскриптов со значимыми изменениями кратности представлен в дополнительной таблице 2, необработанные данные доступны в базе данных GEO под регистрационным номером GSE183629 (удаление и дополнение alr3614 в Anabaena 7120).
Анализы EMSA и TXTL подтверждают функцию репрессора CvkR in vitro
Для проверки регуляции CvkR на идентифицированных ключевых компонентах AnCAST, EMSA и бесклеточные анализы транскрипции-трансляции (TXTL) 9Было выполнено 1378 37 (рис. 5). Очищенный белок CvkR представлял собой димер с молекулярной массой ~ 36,5 кДа по данным анализа эксклюзионной хроматографии (SEC), что примерно вдвое превышает его теоретическую молекулярную массу (~ 17,3 кДа) (рис. 5a). Эта димерная форма в растворе соответствует каноническим регуляторам типа MerR 21,38,39,40,41 . Затем мы создали фрагменты промотора длиной ~80 нт выше или рядом с ранее определенным TSS 28 из cas12k 9.1397 , tnsB , tracrRNA и в качестве отрицательного контроля psbAI (рис. 5b) с 5′-биотиновыми метками и проводили анализы EMSA с рекомбинантным CvkR в различных концентрациях. Результаты подтвердили, что CvkR напрямую связывается с P cas12k , а также с P tnsB , но не с P psbAI (рис. 5c). Удивительно, но между CvkR и P tracr не было связывания (рис. 5с), несмотря на резко измененное содержание транскриптов массива tracrRNA-CRISPR в Δ мутант cvkR in vivo (рис. 4в, г). Чтобы независимо проверить эти результаты, мы клонировали соответствующие промоторы выше репортерного гена deGFP, чтобы проверить, могут ли они управлять транскрипцией и, таким образом, продукцией deGFP в анализе TXTL 37 . Параллельно CvkR экспрессировали из второй плазмиды (рис. 5d). Промоторы P cas12k , P tnsB , P psbAI и P tracr были функциональными и могли управлять экспрессией гена deGFP. В присутствии CvkR продукция deGFP подавлялась в реакциях, содержащих плазмиды с deGFP, управляемые P9.1678 cas12k и P tnsB , но не P tracr и отрицательный контроль P psbAI (рис. 5e, f). Эти результаты хорошо согласуются с анализами гель-сдвига. Мы пришли к выводу, что повышенные уровни мРНК cas12k и tnsB в Δ cvkR были прямым следствием делеции cvkR , тогда как более высокое накопление транскриптов, происходящих от P tracr , должно быть результатом непрямой регуляции CvkR. промоутер cvkR (P cvkR ) не был способен управлять транскрипцией deGFP в системе TXTL (рис. 5e). Следовательно, невозможно проверить, может ли CvkR регулировать собственную транскрипцию. Если перекрывающиеся промоторы cas12k и cvkR были разделены на два фрагмента, мы не наблюдали связывания CvkR с фрагментом P45, содержащим сердцевинную область P cvkR , включая предсказанные элементы -10 и -35 в EMSA. проба. Напротив, сильное связывание CvkR наблюдалось с фрагментом Р43, содержащим сердцевинную область Р9.1678 cas12k (рис. 5б, в). Эти результаты указывают на возможную ауторегуляцию CvkR.
a Оценка молекулярной массы CvkR с помощью эксклюзионной хроматографии (SEC) с колонкой Superdex 75 10/300 GL (GE Healthcare). Калибровочные стандарты указаны. Объем элюирования CvkR составлял ~ 10,86 мл, что соответствует массе ~ 36,5 кДа (вдвое больше расчетной молекулярной массы). mAU, единицы миллиабсорбции. b Последовательности протестированных фрагментов промотора. Зонд Р45 указывает обратную комплементарную последовательность промотора cvkR . Предполагаемые -35 и -10 из промоторных элементов cas12k, tnsB , tracrRNA, psbAI и cvkR показаны синим и зеленым цветом соответственно. TSS были включены и отмечены красным цветом. c Анализы EMSA, показывающие способность связывания CvkR с P cas12k , P tnsB , P tracr , P psbAI и укороченные фрагменты (P43 и P45) P cas12k . Данные репрезентативны из двух или трех независимых экспериментов. Свободный зонд и комплексы комплексов белок-ДНК были помечены как «Свободный» и «Связанный» соответственно. d CvkR экспрессировался из вектора pET28a и был обнаружен вестерн-блоттингом через N-концевую метку 6xHis (верхняя панель), а окрашенная мембрана показана на нижней панели (показан один из трех независимых экспериментов). Соответствующий размер для 6xHis-CvkR равен 19.0,9 кДа. Предварительно окрашенный PageRuler (Thermo Fisher Scientific) использовали в качестве маркера размера. e TXTL assay 37 to compare the promoter activities of cas12k , cvkR, all3630 ( tnsB ) and tracrRNA (P cas12k , P cvkR , P tnsB и P tracr ) для управления экспрессией deGFP в отсутствие или в присутствии CvkR. f Полноразмерная версия P cas12k и фрагмент P43, охватывающий 43 нуклеотида выше TSS cas12k , тестировали в системе TXTL на их способность управлять экспрессией deGFP и опосредовать репрессию при параллельной экспрессии CvkR. P psbAI использовали в качестве отрицательного контроля для промотора, не контролируемого CvkR. Как в e , так и в f , 1 пМ плазмида CvkR экспрессировалась вместе с соответствующими плазмидами р70а (5 нМ) с соответствующими вариантами промотора выше deGFP. Столбики погрешностей показывают стандартные отклонения, рассчитанные для двух технических повторов в одном репрезентативном примере из трех независимых экспериментов. Исходные данные предоставляются в виде файла исходных данных.
Изображение в натуральную величину
Идентификация сайта связывания CvkR
Для дальнейшего определения минимальной необходимой последовательности и мотивов, необходимых для связывания CvkR, P cas12k и P tnsB 9168 проанализированы в анализе футпринтазы IDNB 9168 6а). CvkR специфически защищал 43-нуклеотидный фрагмент P cas12k (P cas12k center ) и 41-нуклеотидный фрагмент P tnsB (P тнсБ центр ). Интересно, что почти идеально консервативная последовательность из 16 нуклеотидов (5′-ATAACATTATGTRTTTT-3′) была общей для защищенных областей P cas12k center и P tnsB center (рис. 6a). Таким образом, эта последовательность лежит в основе потенциального мотива связывания CvkR. Биоинформационный скрининг систем CAST, тесно связанных с AnCAST (дополнительный рисунок 3), выявил в этой последовательности инвертированный повтор длиной 15 нуклеотидов (5′- A TA ACAT 9).2097 T ATGT GT T -3′). Этот потенциальный мотив CvkR широко консервативен в промоторных областях гомологов cas12k и tnsB у цианобактерий (рис. 6b и дополнительная рис. 6).
a Отпечаток ДНКазы I FAM-меченых промоторных последовательностей P cas12k и P tnsB в отсутствие (красные графики) и в присутствии 0,1 мкг Cvk (синий график). Защищенная область заключена в рамку. Отображаются следы секвенирования дидезоксинуклеотидов (зеленый, A; синий, C; черный, G; красный, T), а последовательности ДНК, защищенные CvkR, указаны ниже (для P cas12k ) или выше (для P tnsB ) соответствующие регионы выделены заглавными жирными буквами. Перекрытие последовательностей отмечено вертикальными черными линиями. Пунктирная линия указывает на один другой нуклеотид. Область, прилегающая к CvkR-защищенным областям, отображается строчными буквами. Элементы TSS (красный), −10 (зеленый) и −35 (синий) выделены. b Логотип последовательности мотива CvkR на основе выравнивания последовательностей cas12k и промоторов tnsB 15 систем CAST с генами cvkR (подробности последовательности см. на дополнительном рисунке 6). В логотипах последовательности промоторные области -35 и -10 заштрихованы синим и зеленым цветом, а TSS — коричневым. Защищенные области в анализах следов ДНКазы I выделены синим цветом, а мотив связывания палиндромного CvkR — оранжевым. c Фрагменты промотора дикого типа и мутантные, используемые для анализов EMSA. Зонды P cas12k center и P tnsB center обозначают CvkR-защищенную область промоторов cas12k и tnsB в анализе следов ДНКазы I соответственно. Элементы TSS, -10 и -35 в P cas12k center и P tnsB center обозначены как a . Сайты связывания CvkR обведены розовой рамкой. Перевернутый повтор показан жирным шрифтом и выделен розовыми стрелками, а розовые буквы обозначают мутировавшие сайты. д Взаимодействие между CvkR и вариантами P cas12k проанализировано EMSA. Данные репрезентативны из двух или трех независимых экспериментов. Свободный зонд и комплексы комплексов белок-ДНК помечены как «Свободный» и «Связанный» соответственно. Исходные данные предоставляются в виде файла исходных данных.
Изображение в полный размер
Чтобы подтвердить предполагаемый CvkR-специфический мотив связывания, мы провели анализы EMSA с зондами, в которых внутри палиндрома были заменены разные нуклеотиды (рис. 6c). Эти анализы подтвердили взаимодействие между CvkR и зондами P 9.1678 cas12k center и P tnsB center (рис. 6d), и что CvkR связывается с P wt сходным образом в зависимости от концентрации. Когда обратная комплементарная последовательность в этом мотиве была мутирована (P 15-1 и P 15-2 ) или длина спейсера была изменена (P 15-3 и P 15-4 ), связывание CvkR было почти отменен. Однако простая замена спейсерного нуклеотида с Т на А без изменения длины не дала никакого эффекта (P 15-5 ) (рис. 6в, г). Результаты предоставили убедительные доказательства того, что CvkR связывается с мотивом из 15 нуклеотидов 5′- A nn ACAT n ATGT nn T -3′, и что этот тип связывания CvkR-ДНК широко распространен среди CAST-системы цианобактерий.
Кристаллическая структура показывает, что CvkR представляет собой уникальный регулятор типа MerR
Для исследования функциональности CvkR на структурном уровне была определена высококачественная кристаллическая структура с разрешением 1,6 Å. CvkR состоит из ДНК-связывающего домена классической крылатой спирали-поворота-спирали (wHTH) на N-конце (остатки 1–80), домена димеризации (остатки 81–132) и потенциальной эффекторно-связывающей спирали в С-конце. -концевая область (спираль α7, остатки 133-150), в которой АТФ, используемый в качестве реагента для оптимального формирования кристаллов, был связан в качестве лиганда (рис. 7а). Расположенный на N-конце домен wHTH с топологией α1-α2-W1-α3-W2-α4 является типичной особенностью регуляторов семейства MerR 21,38,39,40,41,42 и, следовательно, указывает на то, что CvkR принадлежит к этому семейству со структурной точки зрения.
a Общая структура комплекса CvkR-АТФ. ДНК-связывающие, димеризационные и предполагаемые эффектор-связывающие С-концевые спиральные домены показаны синим, пурпурным и желтым цветом соответственно. Указаны типичная спираль-поворот-спираль (HTH) и две петли «крыло» W1 и W2 в ДНК-связывающем домене. Лиганд АТФ представлен зеленой палочкой и окрашен в соответствии с типом атома. b Анализ динамического светорассеяния среднего диаметра нативного белка CvkR, измеренного при 25 °C. Данные являются репрезентативными из трех независимых экспериментов. c Мультяшное изображение гомодимера CvkR. Другая субъединица показана белым цветом. Основная часть интерфейса димеризации образована тремя антипараллельными β-тяжами (β2-β1-β3) и одной короткой α-спиралью (α6) из двух субъединиц в димере. Расстояния между двумя взаимодействующими с ДНК спиралями α2/α2’ и между двумя петлями W1 W1/W1’ соответственно измеряют и маркируют. d Интерфейс димеризации в основном состоит из гидрофобных взаимодействий. Ключевые остатки, участвующие в образовании гомодимера, показаны палочками и помечены черным цветом. e , f Сравнение паттернов димеризации белков MerR-типа. Наложения выполняются относительно домена wHTH из одной субъединицы в гомодимерах. Спирали показаны в виде цилиндров для ясности. Наложенные белки: CvkR (эта работа; окраска как у c ), CueR (код PDB 1Q07, две субъединицы окрашены в оранжевый и светло-оранжевый цвета соответственно, эффектор Au показан в красной сфере), GlnR (код PDB 4R4E, две субъединицы окрашены как зеленый и салатовый соответственно). Домены димеризации CueR и GlnR окрашены в красный цвет. г Крупный план взаимодействий для упаковки С-концевой спирали α7. Ключевые остатки показаны палочками, а три солевых мостика отмечены красными пунктирными линиями.
Изображение с полным размером
Члены этого семейства обычно принимают физиологически активную гомодимерную форму 21,22,39,40,43,44,45,46 . Методом динамического рассеяния света (DLS) для CvkR был измерен средний диаметр ~ 5,1 нм (~ 51 Å) (рис. 7b), что согласуется с его сборкой в гомодимер ~ 54 Å в решенной кристаллической структуре (рис. 7c и Дополнительный рис. 7) и результат нашего анализа SEC (рис. 5а). Три антипараллельных β-цепи (β2-β1-β3) и одна короткая α-спираль (α6) вместе с эквивалентными β-цепями и спиралью другой субъединицы составляют основную часть интерфейса димеризации (рис. 7c). Контакты преимущественно гидрофобные (рис. 7г) и закапывают 997,6 Å 2 площадь поверхности (PDBePISA) 47 (дополнительная рис. 7), что намного меньше, чем у канонических белков MerR-типа, например, 1896,8 Å 2 в CueR. Для сравнения, способ димеризации CvkR значительно отличается от регуляторов типа MerR, таких как CueR 39 или MtaN 48 , которые собираются в гомодимеры через двухспиральную антипараллельную спиральную спираль (α5/α5′) (рис. 7e). , а также отличается от необычных регуляторов семейства TnrA/GlnR MerR-типа 39,40 , которые образуют гомодимеры, основанные на N-концевой дополнительной спирали (e-α1/e-α1’) (рис. 7f). Следовательно, паттерн димеризации CvkR уникален среди членов семейства MerR.
Еще одной поразительной особенностью структуры CvkR является то, что С-концевая четырехвитковая α-спираль (α7) упаковывается против своего собственного ДНК-связывающего домена wHTH (α1 и α3) и домена димеризации (от η1 до α5), но не против этих домены своего димерного партнера, как и в других регуляторах типа MerR, таких как CueR или SoxR 39,40 (рис. 7а, д). Три специфических солевых мостика и гидрофобные взаимодействия способствуют точной ориентации спирали α7 (рис. 7e, g).
MerR-подобные белки с распознаваемым C-концевым сенсорным доменом, такие как чувствительные к металлам CueR, MerR и чувствительные к окислительно-восстановительному потенциалу SoxR 39,40,49 , содержат остатки цистеина для эффекторного связывания в консервативных положениях. Однако три цистеина, присутствующие в CvkR, не находятся рядом с потенциальной спиралью α7, связывающей эффектор (рис. 7g и дополнительная рис. 8). Это предполагает, что CvkR может быть одним из тех MerR-подобных белков, у которых отсутствует распознаваемый С-концевой сенсорный домен, такой как HiNmlR из Гемофильная палочка 42 . В нашей решенной структуре реагент, оптимизирующий кристаллы, АТФ был связан рядом с предполагаемым сайтом связывания эффектора посредством стопки π – π, катиона – π и нескольких водородных связей (дополнительная рис. 9a). В то время как неспецифические водородные связи в положениях N1 и N6 аденинового фрагмента вместе с несвязанной трифосфатной группой в предполагаемом сайте связывания эффектора предполагают, что АТФ не является естественным эффектором CvkR, другие производные нуклеозидов могут быть возможными кандидатами (дополнительная рис. . 9б). Действительно, сигнальные молекулы семейства циклических олигонуклеотидов обнаружены в некоторых типах CRISPR-Cas и других защитных системах 50,51 . Однако последующие эксперименты EMSA не показали никакого влияния на связывание ДНК CvkR при добавлении таких лигандов, что указывает на то, что АТФ и его аналоги действительно не являются естественными эффекторами CvkR (дополнительная рис. 10).
Предлагаемая молекулярная основа связывания ДНК CvkR
Для дальнейшего выяснения молекулярной основы связывания ДНК CvkR мы стремились решить структуру комплекса CvkR-промотор, но не достигли лучшего дифракционного разрешения, чем 6 Å, чего недостаточно для определения структуры. Принимая во внимание относительно консервативные характеристики ДНК-связывания домена wHTH среди членов семейства MerR 21,40,42 , мы прибегли к анализу связывания CvkR с ДНК путем структурного выравнивания. CvkR разделяет отсутствие узнаваемой С-концевой сенсорной области и тот факт, что HiNmlR 42 могут нацеливаться на перекрывающиеся промоторы. Поэтому для сравнения был выбран комплекс HiNmlR-ДНК (код PDB 5D8C). Из-за более короткого расстояния между двумя взаимодействующими с ДНК спиралями α2 и α2′ CvkR хорошо совпадал только один домен wHTH (рис. 8a). В соответствии со связыванием ДНК доменов wHTH, на хорошо подобранной стороне спираль α2, богатая положительно заряженными остатками, и крылатая петля W1 отдельно встраиваются в большую и малую бороздки ДНК, в то время как крылатая петля W2 расположена близко к Основа ДНК (рис. 8а, левые панели). На основании сравнения с HiNmlR было высказано предположение, что остатки R20, Q23 и Y24 в α2 и остаток N66 в W2 могут участвовать во взаимодействиях между CvkR и его мишенью (рис. 8a, правая панель) 42 . Более того, положительно заряженные остатки R19, K40 и R42 в спирали α2 и W1, возможно, также участвуют в связывании ДНК, особенно R42, боковая цепь которого одновременно образует водородные связи с α-фосфатной группой АТФ (напоминает фосфат в остове ДНК). в нашей решенной структуре (рис. 8а и дополнительный рис. 9б).
a Верхняя левая панель: наложение CvkR на комплекс HiNmlR-промотор. Две субъединицы гомодимера CvkR окрашены в синий или голубой цвет. Гомодимер HiNmlR и его ДНК-мишень в комплексе HiNmlR-промотор (код PDB 5D8C) отображаются серым и оранжевым цветом соответственно. Помечены три участка, непосредственно участвующие в связывании промоторной ДНК в комплексе HiNmlR-промотор, спираль α2 и две крылатые петли W1 и W2. Нижняя левая панель: изображение электростатической поверхности CvkR с выделением потенциальной электроположительной (синей) ДНК-связывающей поверхности. Правая панель: ключевые взаимодействующие с ДНК остатки, представленные в HiNmlR, показаны палочками и помечены серым цветом. Возможные взаимодействующие с ДНК остатки в CvkR, соответствующие трем взаимодействующим с ДНК областям, отмечены черным цветом. b Анализы EMSA, показывающие связывание вариантов CvkR с P cas12k . Для анализа использовали 33-нуклеотидный зонд P wt . Теги His-SUMO были удалены из всех вариантов CvkR, используемых в анализах EMSA. Свободный зонд и комплексы P wt с белком помечены как «свободный» и «связанный» соответственно. Данные являются репрезентативными из трех независимых экспериментов. c CvkR и мутантные белки экспрессировались в системе TXTL 37 из pET28a(+) или pET28a(+)-SUMO и обнаруживались вестерн-блоттингом по их N-концевой метке 6xHis. Окрашенная мембрана показана ниже. Соответствующие молекулярные массы равны 29.0,9 кДа для 6xHis-SUMO-CvkR WT2 и R42E, в то время как 6xHis-TEV-CvkR WT и 6xHis-TEV-CvkRmut имеют массу 19,9 и 19,5 кДа. d Анализ TXTL для проверки регуляторной способности CvkRmut и R42E по сравнению с CvkR (WT и WT2, как определено на панели C) для подавления флуоресценции deGFP, экспрессируемой промотором cas12k (P cas12k ). Кассету P cas12k -deGFP экспрессировали из плазмиды p70a (5 нМ) в отсутствие или в присутствии 1 пМ pET28a(+), экспрессирующего CvkRmut, R42E, CvkR WT или WT2. Реакции TXTL проводили при 29°C в течение ночи, и флуоресценцию измеряли каждые 10 мин в устройстве для считывания микропланшетов Wallac 1420 Victor2. Планки погрешностей показывают стандартные отклонения, полученные из двух технических повторов в одном репрезентативном примере трех независимых экспериментов. Исходные данные предоставляются в виде файла исходных данных.
Изображение в натуральную величину
Чтобы определить значимость этих остатков для связывания промотора-мишени, были сконструированы два мутантных белка, CvkRmut (R19A-R20A-Q23A-K40A-R42A-N66A) и R42E. Как и в случае с CvkR дикого типа (рис. 5а), оба мутировавших белка появились в форме димера в SEC (дополнительная рис. 11), что позволяет предположить, что они были правильно уложены и пригодны для биохимического анализа. Последующие эксперименты EMSA не показали для обоих белков смещенных полос по сравнению с CvkR дикого типа (рис. 8b). Таким образом, и CvkRmut, и R42E утратили способность специфически связывать P9. 1678 cas12k , прямо подтверждая роль замещенных остатков в связывании ДНК.
Для проверки роли этих остатков в регуляции транскрипции с помощью CvkR были проведены анализы TXTL. Вестерн-блоты показали, что белок CvkR дикого типа стабильно продуцируется как без (WT), так и с меткой SUMO (WT2) в системе TXTL, а также продуцируются мутантные белки CvkR с меткой SUMO (R42E) или 6xHis (CvkRmut). (рис. 8с). Когда CvkRmut и R42E коэкспрессировались с репортерной плазмидой, почти не наблюдалось ингибирующего действия на P9.1678 cas12k по сравнению с контролем CvkR (рис. 8d). Эти наблюдения согласовывались с результатами анализов EMSA и подтверждали нашу модель взаимодействия ДНК с помощью CvkR (рис. 8a, правая панель).
Обсуждение
Системы CAST природного типа V-K до сих пор были обнаружены только у некоторых цианобактерий 11,15 . Их лучшая характеристика представляет фундаментальный интерес, и эти системы открывают большие перспективы для разработки полезных инструментов редактирования генома 9. 1378 52 . Основная функция нативных систем CRISPR-Cas — защита от мобильных генетических элементов. Поэтому мы пришли к выводу, что активность CAST-систем, которые могут транспонироваться в разные сайты генома, должна контролироваться. Здесь мы рассмотрели связь между предполагаемыми регуляторами транскрипции и CAST-системами цианобактерий и обнаружили три различных типа таких репрессоров. Мы предлагаем назвать один из них CvkR для репрессоров Cas-типа V-K, а два других были названы здесь Arc_1 и Arc_2 из-за отсутствия лучшей классификации. Постоянная встречаемость этих регуляторов в 86 из 118 проанализированных систем указывает на функциональную зависимость. Судя по разнообразию регуляторов и их ДНК-связывающих доменов, вполне вероятно, что они приобретались независимо несколько раз.
Было обнаружено, что как cvkR , так и cas12k экспрессируются из безлидерных мРНК. По сравнению с эукариотами и археями частота беслидерных мРНК у бактерий, особенно у грамотрицательных бактерий, довольно низкая 53,54,55 . Анализ dRNA-seq транскриптомов из 14 различных условий роста предоставил доказательства наличия 51 мРНК без лидера в цианобактериях Synechocystis 6803 56 . Даже в хорошо изученном E . coli , глубокое секвенирование обнаружило менее 30 нелидерных мРНК в стандартных условиях роста 57 . Таким образом, биологическая функция беслидерной экспрессии генов в значительной степени еще предстоит установить. мРНК Tn1721 tetR и мРНК λ cI являются наиболее изученными безлидерными мРНК, происходящими из транспозонов и бактериофагов в E . кишечная палочка . Эти безлидерные мРНК были предложены в качестве посредников горизонтального переноса генов, которые контролируются на трансляционном уровне 54 , и это может относиться и к CvkR, особенно потому, что в этом исследовании он был идентифицирован как регулятор системы CAST, которая связана с транспозонами и фагами. Однако реальная значимость трансляции мРНК без лидера в CAST-системах цианобактерий нуждается в дальнейшем изучении.
Затем мы подробно охарактеризовали CvkR, регулятор транскрипции системы AnCAST. Количество мРНК cvkR и белка CvkR в клетках дикого типа было очень низким в тестируемых условиях роста. Однако 9Делеция 1396 cvkR приводила к явно повышенным уровням транскриптов cas12k , tnsB и tracrRNA у мутанта ∆ cvkR . Следовательно, CvkR, вероятно, экспрессируется на низком, но достаточном уровне для выполнения репрессорной функции у Anabaena 7120 дикого типа. Мы выяснили, что CvkR контролирует P cas12k и P tnsB прямо и косвенно количество массива tracrRNA-CRISPR. Мы пришли к выводу, что повышенное накопление tracrRNA может быть результатом защиты посредством связывания с эффектором Cas12k, который присутствует в больших количествах в ∆ cvkR . Все эти результаты указывают на биологическую функцию CvkR как репрессора транскрипции основного модуля AnCAST (т.е. tracrRNA, мРНК cas12k и tnsB ).
Регуляторы типа MerR обычно содержат консервативный N-концевой домен wHTH 22,58 . Они образуют гомодимеры и связывают палиндромный мотив ДНК 49 . Ключевые остатки, непосредственно участвующие во взаимодействии с ДНК, сохраняются у членов семейства MerR, распознающих одни и те же мотивы ДНК, но эти остатки различаются у членов, распознающих разные мотивы ДНК 59 . Путем структурного наложения (рис. 8а) мы предварительно определили остатки CvkR, которые участвуют во взаимодействии с ДНК. Функциональность R42 и дополнительных остатков была подтверждена как в анализе EMSA, так и в анализе TXTL (рис. 8b, d). Основываясь на дальнейшем множественном сравнении последовательностей (дополнительная рис. 12a), мы предлагаем положительно заряженный и высококонсервативный R42 напрямую взаимодействовать с фосфатным остовом ДНК.
Мотивы палиндромной ДНК, распознаваемые белками семейства MerR, обычно расположены в промоторах с удлиненным расстоянием (≥19nt) между элементами -10 и -35. Следовательно, некоторые члены семейства MerR используют механизм искажения ДНК для регуляции экспрессии своих генов-мишеней 22,59 , таких как чувствительные к металлам (т. 45,46 ), редокс-чувствительные (т.е. SoxR 40 и HiNmlR 42 ) и устойчивые к антибиотикам (т.е. BmrR 21 ) регуляторы транскрипции. В этом исследовании мы провели анализ следов ДНКазы I как с P cas12k и P tnsB . Мы получили CvkR-защищенные области с одинаковыми расстояниями и последовательностью длиной 16 нт, общей для двух промоторов. Ориентируясь на эту область, мы в конечном итоге подтвердили мотив CvkR с последовательностью 5′- A nn ACAT n ATGT nn T -3 ‘с помощью серии анализов EMSA и биоинформатического анализа (рис. 6). . При прогнозировании элементов -10 и -35 P cas12k с PromoterHunter 60 , прокладка длиной 21 дюйм, разделяющая блоки -10 (TATATT) и -35 (TTGTAT), явно поддерживалась. Поскольку мотив CvkR в основном расположен внутри этого спейсера, CvkR может модулировать активность P cas12k с использованием механизма MerR-специфического искажения ДНК. Однако для P tnsB мы смогли найти только спейсер -10/-35 из 17 нт (рис. 6b). Как мы показали, связывание CvkR в P tnsB перекрывает TSS P tnsB и частично элемент -10, CvkR должен регулировать экспрессию tnsB иначе, чем cas12k . На самом деле, некоторые регуляторы MerR-типа действительно, как сообщалось, функционируют как репрессоры транскрипции и регулируют экспрессию генов существенно разными способами, такие как BldC 61 , HnoC 62 и McdR 63 . BldC связывает прямые повторы ДНК как кооперативные мультимеры для регуляции экспрессии генов, в то время как HnoC и McdR являются примерами регуляторов, распознающих палиндромные мотивы, перекрывающие TSS и элементы -10. Мы даже заметили спейсер -10/-35 длиной 20 нуклеотидов в промоторе одной из мишеней HnoC, hnoD , что указывает на возможный аналогичный регуляторный механизм вмешательства в ассоциацию холофермента РНКП и инициацию транскрипции , а не на изменение формы промоторной ДНК 62 . McdR, др. репрессор типа MerR, участвующий в контроле клеточного деления, репарации ДНК, устойчивости к лекарствам и др. реакциях на стресс у Mycobacteria, также может репрессировать промоторы-мишени таким образом -63-. Более того, исследованный здесь цианобактериальный CvkR демонстрирует 33% идентичность последовательности с микобактериальным McdR (дополнительная рис. 13a) и обладает структурными особенностями, очень похожими на модель McdR, предсказанную alphafold2 (дополнительная рис. 13b), что подразумевает сходство в механизме регуляции.
В отличие от N-концевого ДНК-связывающего домена регуляторов типа MerR, С-концевые эффекторные связывающие домены не являются консервативными и сильно различаются по последовательности и длине. Множественное выравнивание последовательностей показало, что один из широко консервативных цистеинов в CvkR заменен серином (положение 134, дополнительная фиг. 12a). Поэтому мы создали и протестировали мутантный белок S134C, но в экспериментах SEC, EMSA и TXTL он показал характеристики, очень схожие с белком CvkR дикого типа (дополнительная рис. 12b–d). Структурный анализ также показал, что в CvkR отсутствуют консервативные цистеины для эффекторного связывания на каркасе, построенном доменом димеризации и С-концевым доменом (рис. 7d). Более того, цистеины CvkR распределены структурно дисперсно и расположены далеко от предполагаемого сайта связывания эффектора на основе структурного наложения (дополнительная рис. 8). Эти находки предполагают, что специфическое эффекторное связывание CvkR, вероятно, отличается от обычного цистеин-зависимого способа.
HiNmlR, лишенный распознаваемого С-концевого сенсора, модулирует реакцию на воздействие формальдегида путем модификации двух консервативных остатков цистеина в его домене wHTH (C54 в α3 и C71 в α4) 42 . Два консервативных остатка цистеина C17 и C70 также обнаружены в домене CvkR wHTH, и они взаимодействуют со спиралью α3 в нашей решенной структуре (дополнительный рисунок 8). Кроме того, мы обнаружили, что спираль α3 одновременно взаимодействует с взаимодействующей с ДНК спиралью α2 и С-концевой спиралью α7, что указывает на возможный механизм восприятия структурной перестройки, который может запускаться модификацией остатков цистеина. Кроме того, также возможно, что CvkR связывает эффекторный белок вместо метаболита для осуществления регуляции транскрипции, подобно GlnR, который использует глутаминсинтетазу в качестве эффектора у грамположительных бактерий 64 .
Подводя итог, мы идентифицировали и охарактеризовали CvkR как регулятор системы AnCAST. Структурный анализ CvkR выявил уникальные домены димеризации и связывания лиганда. Эффекторная молекула и функциональность этого гипотетического сигнального входа, воспринимаемого CvkR, являются предметом дальнейших исследований.
Методы
Культуры цианобактерий и конструирование мутантных штаммов
Anabaena 7120 и ее производные выращивали фотоавтотрофно в жидкой среде BG11 или на чашках с агаром при освещении белым светом 30–50 мкмоль фотонов м −2 с − 1 при 30 °C 65 . Что касается контролей в экспериментах, индуцируемых медью, для культивирования использовали прозрачные пластиковые колбы для тканевых культур, для приготовления среды использовали деионизированную воду, а для посевной культуры применяли три ресуспендирования со средой, не содержащей меди. Кроме того, добавляли избыток CuSO 4 (1,0 или 1,25 мкМ), чтобы гарантировать активность P petE в соответствующих экспериментах. В культуру добавляли эритромицин (10 мкг мл −1 ) при необходимости.
Для конструирования Δalr3614 (Δ cvkR ) использовали инструмент редактирования генома CRISPR-Cas12a (Cpf1) вместе с плазмидой pSL2680 (Addgene № 85581) 24 . Пару праймеров alr3614gRNA-1/2 использовали для получения плазмид редактирования гРНК-кассеты, а пары праймеров alr3614KO-1/2 и alr3614KO-3/4 использовали для получения плазмид редактирования гРНК и кассет репарации. Пары праймеров alr3614-3/4 использовали для проверки делеционного генотипа. Внутренний фрагмент длиной 200 п.н. (4365123~4365322) из cvkR был окончательно удален.
Для комплементации Δ мутанта cvkR и/или подтверждения беслидерной экспрессии гена cvkR , три кассеты (P petE _ no 5’UTR _ no 5’UTR , cvFLAG — 9139 P petE _ no 5’UTR — cvkR- S-3xFLAG и P petE — cvkR- S-3xFLAG были клонированы из шаттлового вектора (pRLKR- S-3xFLAG). — плазмида диапазона хозяев RSF1010 путем бесшовной сборки. Праймер пар 59M-F/59M-R1, P petE -F/P petE -R4 и 3614L-F/3614-R использовали для конструирования комплементарной плазмиды для получения Δ cvkR Com-1. Пары праймеров 59M-F/59M-R1, P petE -F/P petE -R5 и 3614S-F1/3614-R использовали для конструирования комплементарной плазмиды для получения Δ cvkR . -2. Пары праймеров 59M-F/59M-R1, P petE -F/P petE -R6 и 3614S-F2/3614-R использовали для конструирования комплементарной плазмиды для получения Δ ЦВКР Ком-3. Каждая комплементарная плазмида была введена в мутант Δ cvkR путем конъюгального переноса 66 . Генотипы мутантов были подтверждены с помощью ПЦР (дополнительный рисунок 4d). Последовательности всех олигонуклеотидов перечислены в дополнительной таблице 3. Все фрагменты ПЦР, плазмиды, созданные в этом исследовании, и области генных мутаций в мутантах были подтверждены секвенированием по Сэнгеру.
Микроматричный анализ
Anabaena 7120 штаммов Δ cvkR и Δ CVKR COM выращивали в 50 мл BG11 без CUSO 4 к OD 750 из 0,8, а экспрессия CVKR для P PETE с 1,25 QM Cus с 1,25 µM CS Cus. Клетки собирали центрифугированием и после ресуспендирования переносили в пробирки с завинчивающимися крышками вместе со стеклянными шариками объемом 250 мкл (диаметром 0,1–0,25 мкм), добавляли 1 мл PGTX 67 и пробирки замораживали в жидком азоте. Разрушение клеток (3 цикла 3 ×6500 об/мин в течение 15 с с 10-секундными перерывами между ними) с помощью гомогенизатора Precellys 24 Dual (Bertin) проводили при охлаждении жидким азотом. Образцы отделяли от гранул и инкубировали 10 мин при 65°С на водяной бане, добавляли один объем хлороформа:изоамилового спирта (24:1) и инкубировали 10–30 мин при комнатной температуре с несколькими циклами встряхивания. . После центрифугирования в течение 3 мин при 3250 г в поворотном роторе, надосадочную жидкость переносили в свежую пробирку и добавляли один объем смеси хлороформ:изоамиловый спирт. Этот шаг был повторен дважды. РНК осаждали добавлением одного объема изопропанола.
Образцы РНК из двух биологических повторов каждый были гибридизированы с микрочипами 8x44K (Agilent ID 062842) в соответствии с опубликованными сведениями о подготовке образцов и гибридизации 68 . Короче говоря, 2 мкг РНК, обработанной ДНКазой, использовали для мечения Cy3 (набор флуоресцентного мечения ULS для массивов Agilent, Kreatech). Гибридизацию на микрочипах проводили с 600 нг Cy3-меченой РНК в течение 17 ч при 65°С. Необработанные данные микрочипов были получены на сканере микрочипов Agilent C, модель G2505C с использованием программного обеспечения Agilent Scan Control and Feature Extraction версии 10.7.3.1. и обработаны с помощью пакета limma R (версия 3.52.4) 69 . Данные были нормализованы с использованием R-реализации в limma 69 , а полученные значения p из t-критерия были скорректированы с использованием метода Бенджамини и Хохберга 70 для контроля уровня ложного обнаружения дифференциального выражения. Пороговое значение |log 2 FC|≥ 1 и значение p ≤ 0,01 указывают на значительное изменение экспрессии генов. Полный набор данных доступен в базе данных GEO под регистрационным номером GSE183629.(Удаление и дополнение alr3614 в Anabaena 7120).
Нозерн-блот-анализ мутантов
Двадцать микрограмм тотальной РНК разделяли на гелях с 10% полиакриламид-8,3 M мочевиной и подвергали электроблотингу на нейлоновые мембраны Hybond N+ (Amersham) при 1 мА на см 2 в течение 1 часа. Накопление транскриптов, связанных с CRISPR, анализировали с помощью нозерн-гибридизации с использованием одноцепочечных радиоактивно меченных РНК-зондов, транскрибированных in vitro с матриц, созданных с помощью ПЦР (праймеры см. в дополнительной таблице 3). Радиоактивно меченные зонды были созданы с использованием [α- 32 P]-UTP и набор для транскрипции Maxiscript T7 In vitro (Thermo Fisher Scientific). Гибридизацию проводили в 50% деионизированном формамиде, 7% SDS, 250 мМ NaCl и 120 мМ Na 2 HPO 4 / NaH 2 PO 4 pH 7,2 в течение ночи при 62 °C. Мембраны промывали в буфере 1 (2 × SSC (3 M NaCl, 0,3 M цитрата натрия, pH 7,0), 1% SDS), буфере 2 (1×SSC, 0,5% SDS) и буфере 3 (0,1 × SSC, 0,1 % SDS) каждый в течение 10 мин. Все этапы промывки проводили при 57 °C. Сигналы были обнаружены с помощью лазерного сканера Typhoon FLA 9.500 (GE Healthcare). Интенсивность сигнала мРНК cas12k (обозначена черным треугольником на рис. 4e) определяли с использованием программного обеспечения Quantity One и нормализовали к сигналу 5S рРНК. Для статистической оценки значений p были рассчитаны из трех биологических повторностей с использованием двустороннего критерия Стьюдента t при условии равных дисперсий с использованием надстройки для анализа данных для программного обеспечения Microsoft Excel 2019.
Вестерн-блот-анализ
Сбор клеток цианобактерий и выделение белка проводили в соответствии с установленными протоколами 71 . Суммарные белки, извлеченные из образцов, разделяли с помощью 15% SDS-PAGE по стандартной методике и подвергали электроблотингу на мембраны PVDF (Amersham). Для проверки одинаковой загрузки мембрану окрашивали Ponceau S (0,1% (масса/объем) в 5% уксусной кислоте). После обесцвечивания мембраны PVDF блокировали в 3% обезжиренном молоке-TBST (0,05% Tween-20 в TBS) при комнатной температуре в течение 30 мин. Затем мембраны инкубировали с мышиным моноклональным антителом против DYKDDDDK (метка FLAG) (#HT201, 1:3000, TransGen) в течение 1 ч и трижды промывали TBST (каждый раз по 15 мин). После этого мембраны инкубировали с Goat Anti-Mouse IgG/AP (#K0031G, 1:3000, Solarbio) в течение 1 часа и трижды промывали TBST (каждый раз по 15 минут). Наконец, сигналы были обнаружены с помощью NBT (хлорид нитро-синего тетразолия) и BCIP (п-толуидиновая соль 5-бром-4-хлор-3′-индолифосфата) (# PR1100, Solarbio). Для вестерн-блоттинга образцов TXTL 1 мкл (рис. 5d) или 9мкл (рис. 8c) реакций TXTL наносили на разделительный гель 15% PAA. Белки электроблоттировали на нитроцеллюлозные мембраны с размером пор 0,45 мкм (Amersham). Набор конъюгатов Penta His HRP (Qiagen) использовали при титре 1:2000 для обнаружения метки 6xHis.
Количественная ПЦР в реальном времени
Экстракцию тотальной РНК, удаление геномной ДНК и обратную транскрипцию проводили с использованием набора Bacteria RNA Extraction Kit и набора HiScript III RT Supermix для qPCR (+gDNA wiper) (Vazyme) в соответствии с инструкциями производителя. . SYBR Premix ExTaqTM (Takara, Далянь, Китай) использовали для qRT-PCR, а пороги цикла определяли с помощью системы обнаружения последовательности Roche LightCycler® 480 II (Roche, Шанхай, Китай). rnpB (субъединица B РНКазы P) использовали в качестве внутреннего контроля. Праймеры для alr3614 ( cvkR ) и rnpB перечислены в дополнительной таблице 3. Было проведено три независимых эксперимента, которые показали согласующиеся результаты.
Анализы TXTL
Для тестирования фрагментов промотора в бесклеточной системе транскрипции-трансляции использовался анализ E. coli на основе TXTL 37,72 . Мастер-микс myTXTL Sigma 70 Cell-Free был приобретен у Arbor Biosciences. Включенную плазмиду p70a использовали в качестве матрицы для клонирования последовательностей промоторов P 9.1678 tracr , P cvkR , P cas12k и P psbAI (рис. 5b) в открытой рамке считывания с дестабилизированной усиленной GFP и ее (5G’UTR) с дестабилизированной усиленной GFP. Все ПЦР проводили с использованием полимеразы PCRBio HiFi (PCR Biosystems). Промоторные последовательности были амплифицированы методом ПЦР из геномной ДНК Anabaena 7120 с перекрыванием p70a. Плазмиду p70a также подвергали ПЦР-амплификации.
Белок CvkR (Alr3614) и его мутантные варианты с потенциально сниженной ДНК-связывающей аффинностью (R16A-R20A-Q23A-K40A-R42A-N66A; CvkRmut и R42E) использовали для анализа в анализе TXTL. 9Последовательность 1396 cvkR была амплифицирована с помощью ПЦР из геномной ДНК Anabaena 7120. Соответствующий фрагмент ДНК для CvkRmut был заказан в IDT как gBlocks и субклонирован в pJet1. 2/blunt (Thermo Fisher Scientific), а затем амплифицирован с помощью ПЦР с выступающими частями до рЕТ28а. Плазмида pET28a также подвергалась ПЦР-амплификации. Таким образом, белки CvkR и CvkRmut экспрессировали с IPTG-индуцируемого промотора T7 с N-концевой меткой 6xHis и сайтом TEV для потенциального расщепления метки.
Сборку фрагмента с помощью клонирования AQUA проводили при комнатной температуре в течение 30 мин после превращения в химически компетентный клеток E. coli DH5α для клонирования. Собранные плазмиды выделяли и секвенировали интересующие области (Eurofins Genomics).
Для экспрессии белков, кодируемых на pET28a, в анализе TXTL необходимы РНК-полимераза T7 (RNAP), экспрессируемая в данном случае из p70a, и IPTG. Реакции проводили в двух повторностях в течение ночи при 29°C в общем объеме 5 мкл с 3,75 мкл мастер-микса TXTL, 1 мМ IPTG, 0,5 нМ p70a_T7_RNAP, 1 пМ pET28a_CvkR (или мутанты: CvkRmut, R42E, S134C) и 5 нМ p70a_FPpromoter_deGFP. . Флуоресценцию deGFP измеряли каждые 10 мин (возбуждение 485 нм, эмиссионный фильтр 535 нм) в устройстве для чтения планшетов (Wallac 1420 Victor2 для чтения микропланшетов от Perkin Elmer).
Гетерологическая экспрессия белка и очистка
Для гетерологичной экспрессии CvkR в E. coli кодирующую белок последовательность CvkR амплифицировали с помощью ПЦР из геномной ДНК Anabaena 7120 и клонировали в вектор pET28a-SUMO с использованием BamHI. и сайты рестрикции XhoI для получения экспрессионной плазмиды pET28a-SUMO_CvkR, которая экспрессирует CvkR с расщепляемой Ulp1 N-концевой слитой меткой 6xHis-SUMO. Плазмиды pET28a-SUMO_CvkR-R42E и -S134C, используемые для получения мутантных белков R42E и S134C, конструировали методом точечной мутации с использованием плазмиды pET28a-SUMO_CvkR в качестве матрицы. Плазмида pET28a-SUMO_CvkRmut, используемая для получения CvkRmut (R16A-R20A-Q23A-K40A-R42A-N66A), была сконструирована путем бесшовной сборки с использованием pET28a-SUMO_CvkR в качестве основы и упорядоченного фрагмента ДНК для CvkRmut в анализе TXTL в качестве вставной кассеты. Затем плазмиды с проверенной последовательностью трансформировали в E. coli BL21(DE3) для экспрессии белка.
Экспрессионные штаммы выращивали в среде LB приблизительно до OD 600 ~ 0,6 при 37°C и индуцировали при 25°C в течение ночи с помощью 0,4 мМ IPTG (изопропил β-D-тиогалактопиранозид). После индукции клетки собирали центрифугированием и лизировали в лизирующем буфере А (20 мМ трис-HCl, pH 8,0, 300 мМ NaCl, 10 мМ имидазол, 2 мМ β-меркаптоэтанол, 20 мкг мл -1 ДНКазы I) с использованием высокой — гомогенизатор давления или ультразвуковой разрушитель клеток. После центрифугирования при 20 000 × г в течение 50 мин, супернатанты наносили на колонку Ni Sepharose 6 FF (GE Healthcare), промывали лизирующим буфером А, содержащим 40 мМ имидазол, и элюировали 20 мМ трис-HCl (pH 8,8), 300 мМ имидазола, и 2 мМ β-меркаптоэтанола. Элюированные фракции обрабатывали протеазой Ulp1 в течение ночи при 4 °C и очищали с помощью колонки HiTrap Heparin HP (GE Healthcare) для удаления слитой метки 6xHis-SUMO и примесей. Впоследствии фракции целевого белка с удаленными метками дополнительно очищали с помощью Superdex 75 10/300 GL (GE Healthcare) с 10 мМ Tris-HCl (pH 8,8), 100 мМ NaCl и 2 мМ β-меркаптоэтанола. Наконец, целевые белки собирали, концентрировали и хранили в том же буфере. Все мутанты очищали по тому же протоколу, что и для получения белка CvkR дикого типа.
Меченый селенометионином (Se-Met) белок CvkR подвергали сверхэкспрессии и очищали с использованием тех же процедур, которые описаны выше; однако среду заменяли средой М9, и перед индукцией в середине логарифмической фазы добавляли семь незаменимых аминокислот. Для предотвращения окисления атомов селена к конечной фракции элюирования, содержащей Se-Met CvkR, добавляли 5 мМ β-меркаптоэтанола.
Аналитическая гель-фильтрация
Эксклюзионная хроматография была проведена для определения молекулярной массы CvkR и соответствующих мутантных белков в растворе с использованием колонки Superdex 75 10/300 GL, соединенной с системой ÄKTA Pure (GE Healthcare). Колонку калибровали с помощью калибровочного набора для гель-фильтрации LMW (Low Molecular Weight) (GE Healthcare) в буфере, содержащем 10 мМ Tris-HCl (pH 8,8), 100 мМ NaCl, 2 мМ β-меркаптоэтанола. Около 500 мкл белков (~0,6 мг мл -1 ) загружали в систему с помощью петли на 500 мкл. Калибровочные кривые использовали для расчета олигомерного состояния немеченых CvkR, R42E, S134C и CvkRmut в соответствии с их объемами элюции. Калибровочная кривая, основанная на молекулярных маркерах (Cytiva) (кональбумин: 75 кДа, овальбумин: 44 кДа, карбоангидраза: 29 кДа, рибонуклеаза A: 13,7 кДа, апротинин: 6,5 кДа) представляет собой lgMr = −1,7621Ve 8+ 4. 2 = 0,9841, Ve: объем элюирования, Mr: молекулярная масса).
Анализ динамического светорассеяния
Анализ DLS проводили с использованием прибора для считывания планшетов DynaPro99 DLS (Wyatt), оснащенного лазерным источником с длиной волны 830 нм. Белок CvkR использовали в конечной концентрации 50 мкМ в буфере, содержащем 10 мМ Tris-HCl (pH 8,8), 100 мМ NaCl, и измерения проводили в течение 10 мин при 25 °C. Программное обеспечение Wyatt Dynamics (версия 7.0) использовалось для планирования сбора данных и расчета гидродинамического радиуса.
Анализы EMSA
Двухцепочечные ДНК-зонды для EMSA были получены путем отжига 5′-биотин-меченых олигонуклеотидов и их комплементарных цепей (Tsingke Biotechnology Co., Ltd.) (дополнительная таблица 3) в эквимолярных концентрациях (10 мкМ ) с термоциклером (94 °С в течение 5 мин, затем со снижением температуры на 1 °С в минуту до 25 °С). Анализы EMSA проводили с использованием хемилюминесцентного набора EMSA (Beyotime, Китай) в соответствии с протоколом производителя. Каждый двухцепочечный ДНК-зонд с концентрацией 2 нМ и различные количества представляющих интерес CvkR или мутантных белков смешивали с буфером для связывания EMSA до конечного объема 10 мкл и инкубировали при 25 °С в течение 30 мин. Буфер для связывания содержит 10 мМ Tris-HCl (pH 8,0), 25 мМ MgCl 2 , 50 мМ NaCl, 1 мМ дитиотреитола, 1 мМ ЭДТА, 50 мкг 9 мл 9.1378 -1 поли(dI-dC), 0,01% Nonidet P-40 и 10% глицерина. После связывания образцы подвергали электрофорезу в 6% нативном полиакриламидном геле с 0,5x трис-борат-ЭДТА (ТВЕ) буфером с последующим электропереносом на нейлоновую мембрану и сшивкой с помощью ручной УФ-лампы. Полосы обнаруживали с помощью набора BeyoECL Plus (Beyotime).
Анализы футпринтинга ДНКазы I
Анализы футпринтинга ДНКазы I проводились аналогично предыдущим исследованиям 73 . В частности, зонды, меченные FAM ~300 н., которые покрывают промоторные области cas12k или tnsB были клонированы в вектор pUCm-T и ПЦР амплифицированы с 2-кратным премиксом ДНК-полимеразы TOLO HIFI (TOLO Biotech, Шанхай) с использованием пар праймеров P3613-F(FAM)/P3613-R или M13F(FAM) /M13R (дополнительная таблица 3) и очищены с помощью геля Wizard® SV и системы очистки ПЦР (Promega, США). Реакции связывания проводили в общем объеме 40 мкл, содержащем 50 мМ Tris-HCl, pH 8,0, 100 мМ KCl, 2,5 мМ MgCl 2 , 0,2 мМ DTT, 10% глицерина, 2 мкг ДНК спермы лосося, 300 нг зондов и 0,1 мкг белка CvkR при комнатной температуре в течение 30 мин. После обработки ДНКазой I (Promega), экстракции фенолом/хлороформом и осаждения этанолом продукты растворяли в 30 мкл воды MilliQ. Подготовка лестницы ДНК, электрофорез и анализ данных проводились в соответствии с протоколом исх. 73 , за исключением того, что использовался стандарт размера GeneScan-LIZ600 (Applied Biosystems).
Филогенетический анализ CvkR и Cas12k
Идентифицированные белки CvkR и Cas12k сравнивали друг с другом и анализировали на наличие альтернативных стартовых положений для исправления потенциально неверных аннотаций, как в случае Alr3614 (BAB75313.1) и All3613 (BAB75312.1) ). Удлиненные N-концевые участки, не имеющие сходства с гомологами, кодируемыми другими генами cvkR и cas12k , были удалены из анализа. Затем последовательности выравнивали с помощью M-coffee 9.1378 74,75 и далее проанализировано алгоритмом BEAST(v 1.10.4) 76 . Филогенетический анализ был рассчитан с использованием строгой модели часов и процесса видообразования при рождении-смерти (Yule) в качестве априорного дерева с использованием стандартных параметров (clock. rate: фиксированное значение = 1; treeModel.rootHeight: с использованием априорного дерева [0, бесконечность] и yule.birthRate: LogNormal [1, 1.5], initial = 2), а также модель подстановки Blosum62. Цепь MCMC проверялась на CvkR каждые 1000 шагов в течение 1 миллиона поколений 9.1378 77,78,79 и каждые 1000 шагов через 10 млн поколений 77,78,79 для анализа Cas12k. Чтобы обеспечить разумный эффективный размер выборки (ESS; пороговое значение 200), результаты были проанализированы с помощью инструмента Tracer v 1.7.2 80 , и первые 10% деревьев были исключены из анализа. Затем деревья были сгенерированы с помощью инструмента аннотации деревьев (из пакета BEAST).
Кристаллизация, сбор данных и определение структуры белка CvkR
С помощью метода диффузии паров висящих капель, как нативные, так и селенометионин-замещенные (Se-Met) кристаллы CvkR появлялись при 18 °C в реагенте для кристаллизации, содержащем 0,1 M цитрата фосфата (pH 4,4~4,6) с 15%~20% PEG300 . Однако кристаллы дифракционного качества были получены только при добавлении 10 мМ АТФ в каплю кристаллического экрана, которая состояла из белка 1:1 (об./об.) в количестве 15 мг/мл -1 и реагента для кристаллизации лунок. Перед мгновенным охлаждением в жидком азоте кристаллы подвергали криоконсервации с использованием реагента для кристаллизации с добавлением 15% ПЭГ400.
Данные дифракции были собраны на длинах волн 0,97930 Å или 0,97918 Å на Шанхайской установке синхротронного излучения, линии пучка BL17U1 и BL18U1, в потоке азота при 100 K. Индексация данных, интеграция и масштабирование для собственного CvkR проводились с использованием программного обеспечения HKL3000 (версия 721.3) 81 . Данные рентгеновской дифракции SAD были обработаны трубопроводом Aquarium 82 . Одиннадцать сайтов с селеном были обнаружены с помощью HySS, реализованного в AutoSol/Phenix (версия 1.17.1-3660). Полученная структура основной цепи использовалась в качестве начальной модели поиска молекулярной замены с помощью Phaser/Phenix (версия 1. 17.1-3660) для определения нативной структуры CvkR 9.1378 83 . Уточнение структуры выполняли итеративно с использованием программ Phenix и Coot (версия 0.9.8.1 EL) 84,85 . Статистика обработки данных и уточнения структуры показана в дополнительной таблице 4. Координаты были депонированы в Банк данных белков с идентификатором PDB 7XN2. Рисунки были подготовлены с использованием PyMOL (версия 2.4.0) 86 .
Сводка отчета
Дополнительная информация о дизайне исследования доступна в Резюме отчета Nature Portfolio, связанном с этой статьей.
Доступность данных
Исходные данные предоставляются в виде файла исходных данных. Полные наборы данных транскриптома для WT и мутантов Δ cvkR и Δ cvkR Com доступны в базе данных GEO под регистрационным номером GSE183629 (удаление и дополнение alr3614 в Anabaena 7120). Доступ к структурным данным можно получить в Protein Data Bank под регистрационным номером PDB 7XN2. Другими структурами PDB, упомянутыми в этом исследовании, являются 1Q07 (структура CueR), 4R4E (структура GlnR-ДНК) и 5D8C (структура HiNmlR-ДНК). AlphaFold2 предсказал McdR (NCBI: ABK74795.1) [https://alphafold.ebi.ac.uk/entry/A0QYF9]. Исходные данные приводятся вместе с настоящей статьей.
Ссылки
Bhaya, D., Davison, M. & Barrangou, R. Системы CRISPR-Cas у бактерий и архей: универсальные малые РНК для адаптивной защиты и регуляции. год. Преподобный Жене. 45 , 273–297 (2011).
Артикул КАС пабмед Google Scholar
Grissa, I., Vergnaud, G. & Pourcel, C. База данных CRISPRdb и инструменты для отображения CRISPR и создания словарей спейсеров и повторов. БМК Биоинформ. 8 , 172 (2007).
Артикул Google Scholar
Ланге, С. Дж., Алхнбаши, О. С., Роуз, Д., Уилл, С. и Баккофен, Р. CRISPRmap: автоматизированная классификация сохранения повторов в прокариотических адаптивных иммунных системах. Рез. нуклеиновых кислот. 41 , 8034–8044 (2013).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Макарова К.С. и др. Обновленная эволюционная классификация систем CRISPR-Cas. Нац. Преподобный Микробиолог. 13 , 722–736 (2015).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Нотт, Г. Дж. и Дудна, Дж. А. CRISPR-Cas определяет будущее генной инженерии. Наука 361 , 866–869 (2018).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Идальго-Кантабрана, К., Гох, Ю. Дж. и Баррангу, Р. Характеристика и перепрофилирование систем CRISPR-Cas типа I и типа II в бактериях. Дж. Мол. биол. 431 , 21–33 (2019).
Артикул КАС пабмед Google Scholar
Terns, M. P. Технологии на основе CRISPR: влияние систем нацеливания на РНК. Мол. Cell 72 , 404–412 (2018).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Макарова К.С. и др. Эволюционная классификация систем CRISPR-Cas: взрыв класса 2 и производные варианты. Нац. Преподобный Микробиолог. 18 , 67–83 (2020).
Артикул КАС пабмед Google Scholar
Strecker, J. et al. Вставка ДНК под управлением РНК с CRISPR-ассоциированными транспозазами. Наука 365 , 48–53 (2019).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Klompe, S.E., Vo, PLH, Halpin-Healy, T.S. & Sternberg, S.H. Системы CRISPR-Cas, кодируемые транспозонами, обеспечивают прямую интеграцию ДНК под управлением РНК. Природа 571 , 219–225 (2019).
Артикул КАС пабмед Google Scholar
Klompe, S.E. et al. Эволюционное и механистическое разнообразие CRISPR-ассоциированных транспозонов типа I-F. Мол. Cell 82 , 616–628.e5 (2022).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Хоу, С. и др. Системы CRISPR-Cas у многоклеточных цианобактерий. РНК Биол. 16 , 518–529 (2019).
Артикул пабмед Google Scholar
Schmitz, M., Querques, I., Oberli, S., Chanez, C. & Jinek, M. Структурная основа для сборки CRISPR-ассоциированного транспозонного комплекса типа V. Сотовый 185 4999–5010.
https://doi.org/10.1016/j.cell.2022.11.009 (2022).Парк, Ж.-Ю. и другие. Структурная основа для выбора сайта-мишени в системах транспозиции ДНК, управляемых РНК. Наука 373 , 768–773 (2021).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Chen, W. et al. Целевой генетический скрининг бактерий с транспозазой, управляемой Cas12k. Cell Rep. 36 , 109635 (2021).
Артикул КАС пабмед Google Scholar
Vo, P.L.H. et al. CRISPR РНК-управляемые интегразы для высокоэффективной мультиплексной инженерии бактериального генома. Нац. Биотехнолог. 39 , 480–489 (2021).
Артикул КАС пабмед Google Scholar
Рубин Б.
Е. и др. Видо- и сайт-специфичное редактирование генома в сложных бактериальных сообществах. Нац. микробиол. 22 , 34–47 (2022).Google Scholar
Heldwein, E.E. & Brennan, R.G. Кристаллическая структура активатора транскрипции BmrR, связанного с ДНК и лекарственным средством. Природа 409 , 378–382 (2001).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед Google Scholar
Браун Н.Л., Стоянов Дж.В., Кидд С.П. и Хобман Дж.Л. Семейство регуляторов транскрипции MerR. FEMS Microbiol. 27 , 145–163 (2003).
Артикул КАС пабмед Google Scholar
Филипс, С.Дж., Канализо-Эрнандес, М., Йилдирим, И., Шац, Г.К. и Мондрагон, А. Аллостерическая регуляция транскрипции посредством изменений в общей топологии основного промотора.
Наука 349 , 877–881 (2015).Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Reimann, V. et al. Специфичность и функциональная координация между двумя эндонуклеазами созревания Cas6 у Anabaena sp. PCC 7120 распределяет потерянные массивы CRISPR по трем группам. РНК Биол. 17 , 1442–1453 (2020).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Querques, I., Schmitz, M., Oberli, S., Chanez, C. & Jinek, M. Выбор и ремоделирование целевого сайта с помощью систем CRISPR-транспозонов типа V. Природа 599 , 497–502 (2021).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Xiao, R. et al. Структурные основы распознавания ДНК-мишени с помощью CRISPR-Cas12k для РНК-управляемой транспозиции ДНК.
Мол. Ячейка 81 , 4457–4466 (2021).Артикул КАС пабмед ПабМед Центральный Google Scholar
Митрофанов А., Циманн М., Альхнбаши О.С., Хесс В.Р. и Бакофен Р. CRISPRtracrRNA: надежный подход к обнаружению CRISPR tracrRNA. Биоинформатика 38 , ii42–ii48 (2022).
Артикул пабмед ПабМед Центральный Google Scholar
Митшке, Дж., Виоке, А., Хаас, Ф., Хесс, В. Р. и Муро-Пастор, А. М. Динамика выбора сайта начала транскрипции во время дифференцировки клеток, вызванной азотным стрессом, у Anabaena sp. ПКС7120. Проц. Натл акад. науч. 108 , 20130–20135 (2011).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Vo, P.L.H., Acree, C.
, Smith, M.L. & Sternberg, S.H. Беспристрастное профилирование продуктов транспозиции, управляемой CRISPR РНК, путем секвенирования с длинным считыванием. Моб. ДНК 12 , 13 (2021).Артикул КАС пабмед ПабМед Центральный Google Scholar
Альтшул, С. Ф., Гиш, В., Миллер, В., Майерс, Э. В. и Липман, Д. Дж. Базовый инструмент локального поиска выравнивания. Дж. Мол. биол. 215 , 403–410 (1990).
Артикул КАС пабмед Google Scholar
Лу, С. и др. CDD/SPARCLE: сохраненная база данных доменов в 2020 г. Рез. нуклеиновых кислот. 48 , Д265–Д268 (2020).
Артикул КАС пабмед Google Scholar
Soding, J. Определение гомологии белков путем сравнения HMM-HMM. Биоинформатика 21 , 951–960 (2005).
Артикул пабмед Google Scholar
Шрайтер, Э. Р. и Дреннан, К. Л. Факторы транскрипции лента-спираль-спираль: вариации на тему. Нац. Преподобный Микробиолог. 5 , 710–720 (2007).
Артикул КАС пабмед Google Scholar
Шильдбах, Дж. Ф., Карзай, А. В., Рауманн, Б. Е. и Зауэр, Р. Т. Происхождение специфичности связывания ДНК: роль белковых контактов с остовом ДНК. Проц. Натл акад. науч. 96 , 811–817 (1999).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Бренес-Альварес, М. и др. Элементы специфичного для гетероцист транскриптома, обнаруженные с помощью анализа коэкспрессии у Nostoc sp. PCC 7120. Окружающая среда. микробиол. 21 , 2544–2558 (2019).
Артикул пабмед Google Scholar
Сантамария-Гомес, Дж. и др. Роль криптического оперона гена тРНК в выживании в условиях трансляционного стресса. Рез. нуклеиновых кислот. 49 , 8757–8776 (2021).
Артикул пабмед ПабМед Центральный Google Scholar
Максвелл, К.С., Якобсен, Т., Маршалл, Р., Нуаро, В. и Бейзел, К.Л. Подробный бесклеточный анализ на основе транскрипции-трансляции для расшифровки мотивов CRISPR, смежных с протоспейсером. Методы. 143 , 48–57 (2018).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Годси, М. Х., Баранова, Н. Н., Нейфах, А. А. и Бреннан, Р. Г. Кристаллическая структура MtaN, глобальный активатор гена-переносчика лекарств. J. Biol. хим. 276 , 47178–47184 (2001).
Артикул КАС пабмед Google Scholar
Чангела, А. и др. Молекулярная основа селективности ионов металлов и зептомолярной чувствительности с помощью CueR. Наука 301 , 1383–1387 (2003).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед Google Scholar
Ватанабе С., Кита А., Кобаяши К. и Мики К. Кристаллическая структура [2Fe-2S] датчика окислительного стресса SoxR, связанного с ДНК. Проц. Натл. акад. науч. США 105 , 4121–4126 (2008 г.).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Ван, Ф. и др. БРр от Pseudomonas aeruginosa является рецептором циклического ди-ГМФ и пиоцианина. Нац. коммун. 9 , 2563 (2018).
Артикул ОБЪЯВЛЕНИЯ пабмед ПабМед Центральный Google Scholar
O’Halloran, T. & Walsh, C. Металлорегуляторный ДНК-связывающий белок, кодируемый геном mer : выделение и характеристика. Наука 235 , 211–214 (1987).
Артикул ОБЪЯВЛЕНИЯ пабмед Google Scholar
Brocklehurst, K.R. et al. ZntR представляет собой Zn(II)-чувствительный MerR-подобный регулятор транскрипции zntA в Escherichia coli . Мол. микробиол. 31 , 893–902 (1999).
Артикул КАС пабмед Google Scholar
Фанг, К. и др. CueR активирует транскрипцию посредством механизма искажения ДНК. Нац. хим. биол. 17 , 57–64 (2021).
Артикул КАС пабмед Google Scholar
Криссинел, Э. и Хенрик, К. Вывод макромолекулярных ансамблей из кристаллического состояния. Дж. Мол. биол. 372 , 774–797 (2007).
Артикул КАС пабмед Google Scholar
Ньюберри, К. Дж. и Бреннан, Р. Г. Структурный механизм активации транскрипции с помощью активации множественного переносчика лекарств члена семейства MerR, N-конец.
J. Biol. хим. 279 , 20356–20362 (2004 г.).Артикул КАС пабмед Google Scholar
Чанг, К.-К., Линь, Л.-Ю., Цзоу, X.-В., Хуан, К.-К. и Чан, Н.-Л. Структурные основы опосредованного ртутью(II) конформационного переключения двухфункционального регулятора транскрипции MerR. Рез. нуклеиновых кислот. 43 , 7612–7623 (2015).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Athukoralage, J. S. et al. Динамическое взаимодействие ферментов хозяина и вируса в передаче сигналов циклических нуклеотидов, опосредованной CRISPR типа III. eLife 9 , e55852 (2020).
Артикул пабмед ПабМед Центральный Google Scholar
Миллман А., Меламед С., Амитай Г. и Сорек Р. Разнообразие и классификация антифаговых сигнальных систем на основе циклических олигонуклеотидов.
Нац. микробиол. 5 , 1608–1615 (2020).Артикул КАС пабмед ПабМед Центральный Google Scholar
Tou, C.J., Orr, B. & Kleinstiver, B.P. Точная вставка ДНК путем вырезания и вставки с использованием сконструированных CRISPR-ассоциированных транспозаз типа V-K. Нац. Биотехнолог . 1–12 https://doi.org/10.1038/s41587-022-01574-x (2023).
Лейва, Л. Э. и Кац, А. Регуляция трансляции мРНК без лидера у бактерий. Микроорганизмы 10 , 723 (2022).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Moll, I., Grill, S., Gualerzi, C. O. & Bläsi, U. Безлидерные мРНК в бактериях: неожиданности в рибосомном рекрутировании и контроле трансляции. мол. микробиол. 43 , 239–246 (2002).
Артикул КАС пабмед Google Scholar
Копф, М. и др. Сравнительный анализ первичного транскриптома Synechocystis sp. PCC 6803. DNA Res. 21 , 527–539 (2014).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Мендоза-Варгас, А. и др. Полногеномная идентификация сайтов начала транскрипции, промоторов и сайтов связывания факторов транскрипции в E. coli . PloS Один 4 , e7526 (2009 г.).
Артикул ОБЪЯВЛЕНИЯ пабмед ПабМед Центральный Google Scholar
Ван, Д. и др. Структурный анализ Hg(II)-регуляторного белка Tn501 MerR из Pseudomonas aeruginosa .
науч. 6 , 33391 (2016).Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Fang, C. & Zhang, Y. Бактериальные регуляторы транскрипции семейства MerR: активация путем искажения. Акта Биохим. Биофиз. Грех. 54 , 25–36 (2022).
Артикул КАС пабмед Google Scholar
Клукар Л., Стано М. и Хайдук М., phiSITE: база данных регуляции генов бактериофагов. Рез. нуклеиновых кислот. 38 , Д366–Д370 (2010).
Bush, M.J., Chandra, G., Al-Bassam, M.M., Findlay, K.C. & Buttner, M.J. BldC задерживает начало развития, чтобы обеспечить устойчивый период вегетативного роста у Streptomyces venezuelae . mBio 10 , e02812–18 (2019).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Zhou, W. et al. Регуляторная петля обратной связи, содержащая McdR и WhiB2, контролирует деление клеток и восстановление ДНК у микобактерий. мбио 13 , 14 (2022).
Артикул Google Scholar
Трэвис Б.А. и др. Молекулярное рассечение схемы регуляции азота глутаминсинтетаза-GlnR у грамположительных бактерий. Нац. коммун. 13 , 3793 (2022).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Эльхай, Дж. и Волк, С.П. Конъюгальный перенос ДНК в цианобактерии. Методы Фермент. 167 , 747–754 (1988).
Артикул КАС пабмед Google Scholar
Пинто, Ф., Таппер, А., Сонтхейм, В. и Линдблад, П. Анализ существующих и альтернативных методов выделения РНК на основе фенола для цианобактерий. ВМС мол. биол. 10 , 79 (2009).
Артикул пабмед ПабМед Центральный Google Scholar
Восс, Б. и Хесс, В. Р. Идентификация бактериальных некодирующих РНК с помощью дополнительных подходов.
Справочник по биохимии РНК 787–800 (John Wiley & Sons, Ltd, 2014). https://doi.org/10.1002/9783527647064.ch44.Ritchie, M.E. et al. limma обеспечивает анализ дифференциальной экспрессии для секвенирования РНК и исследований микрочипов. Рез. нуклеиновых кислот. 43 , e47 (2015).
Артикул пабмед ПабМед Центральный Google Scholar
Бенджамини Ю. и Хохберг Ю. Контроль уровня ложных открытий: практичный и мощный подход к множественному тестированию. JR Stat. соц. сер. Б Методол. 57 , 289–300 (1995).
MathSciNet МАТЕМАТИКА Google Scholar
Zhu, T., Xie, X., Li, Z., Tan, X. и Lu, X. Повышение фотосинтетического производства этилена в генно-инженерном Synechocystis sp. PCC 6803. Зеленый. хим. 17 , 421–434 (2015).
Артикул КАС Google Scholar
Shin, J. & Noireaux, V. Набор инструментов для бесклеточной экспрессии E. coli : применение к синтетическим генным цепям и искусственным клеткам. ACS Синтез. биол. 1 , 29–41 (2012).
Артикул КАС пабмед Google Scholar
Ван, Ю., Сен, X.-Ф., Чжао, Г.-П. & Wang, J. Характеристика нового связывающего бокса GlnR в промоторе amtB в Streptomyces coelicolor позволила сделать вывод о механизме конкурентного связывания PhoP/GlnR для регуляции транскрипции amtB . J. Бактериол. 194 , 5237–5244 (2012).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Ди Томмазо, П. и др. T-Coffee: веб-сервер для множественного выравнивания последовательностей белков и РНК с использованием структурной информации и расширения гомологии.
Рез. нуклеиновых кислот. 39 , W13–W17 (2011).Артикул пабмед ПабМед Центральный Google Scholar
Нотредам К., Хиггинс Д. Г. и Херинга Дж. Т-кофе: новый метод быстрого и точного выравнивания множественных последовательностей. Дж. Мол. биол. 302 , 205–217 (2000).
Артикул КАС пабмед Google Scholar
Сучард, М. А. и др. Интеграция байесовских филогенетических и филодинамических данных с использованием BEAST 1.10. Эволюция вируса. 4 , vey016 (2018).
Артикул пабмед ПабМед Центральный Google Scholar
Гернхард Т. Условно реконструированный процесс. Ж. Теор. биол. 253 , 769–778 (2008).
Артикул ОБЪЯВЛЕНИЯ MathSciNet пабмед МАТЕМАТИКА Google Scholar
Yule, U. Математическая теория эволюции, основанная на выводах доктора JC Willis, FRS Philos. Транс. Р. Соц. Лонд. сер. B 213 , 21–87 (1925).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
Рамбо, А., Драммонд, А.Дж., Се, Д., Баеле, Г. и Сучард, М.А. Апостериорное обобщение в байесовской филогенетике с использованием индикатора 1.7. Сист. биол. 67 , 901–904 (2018).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Майнор, В., Цимборовски, М., Отвиновски, З.
и Хрущ, М. HKL-3000: интеграция обработки данных и структурного решения – от дифракционных изображений до исходной модели за считанные минуты. Acta Кристаллогр. Д. биол. Кристаллогр. 62 , 859–866 (2006).Артикул пабмед Google Scholar
Ю. Ф. и др. Аквариум: автоматическая система обработки данных и управления экспериментальной информацией для линий биологической макромолекулярной кристаллографии. J. Appl. Кристаллогр. 52 , 472–477 (2019).
Артикул КАС Google Scholar
McCoy, A.J. et al. Кристаллографическая программа Phaser. J. Appl. Кристаллогр. 40 , 658–674 (2007).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Адамс, П. Д. и др. PHENIX: комплексная система на основе Python для решения макромолекулярной структуры.
Acta Кристаллогр. Д. биол. Кристаллогр. 66 , 213–221 (2010).Артикул КАС пабмед ПабМед Центральный Google Scholar
Эмсли П., Локамп Б., Скотт В. Г. и Коутан К. Особенности и развитие Coot. Acta Кристаллогр. Д. биол. Кристаллогр. 66 , 486–501 (2010).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Шредингер. ООО. Система молекулярной графики PyMOL, версия 2.0. (2015).
Katoh, K. & Standley, D.M. MAFFT Multiple Sequence Alignment Software, версия 7: улучшения производительности и удобства использования. Мол. биол. Эвол. 30 , 772–780 (2013).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Оконечников К.
, Голосова О. и Фурсов М., команда UGENE. Юнипро ЮГЕНЭ: единый инструментарий биоинформатики. Биоинформа 28 , 1166–1167 (2012).Артикул КАС Google Scholar
Янсен Р., ван Эмбден Дж. Д. А., Гаастра В. и Шоулс Л. М. Идентификация генов, связанных с повторами ДНК у прокариот. Мол. микробиол. 43 , 1565–1575 (2002).
Артикул КАС пабмед Google Scholar
Хохштрассер, М. Л. и Дудна, Дж. А. Вкратце: структура и функция эндорибонуклеазы, связанной с CRISPR. Тенденции биохим. науч. 40 , 58–66 (2015).
Артикул КАС пабмед Google Scholar
Сайто, М. и др. Двойной режим CRISPR-ассоциированного самонаведения транспозонов. Cell 184 , 2441–2453.e18 (2021).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Couñago, R. M. et al. Структурная основа тиоловой регуляции детоксикации формальдегида в H. influenzae с помощью регулятора MerR без сенсорной области. Рез. нуклеиновых кислот. 44 , 6981–6993 (2016).
Артикул пабмед ПабМед Центральный Google Scholar
Fang, C. et al. Бактериальный регулятор множественной лекарственной устойчивости BmrR искажает промоторную ДНК, чтобы активировать транскрипцию. Нац. коммун. 11 , 6284 (2020).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Чжэн, С., Ху, Г.-К., Ше, З.-С. & Zhu, H. Гены без лидера у бактерий: ключ к эволюции механизмов инициации трансляции у прокариот. BMC Геном. 12 , 361 (2011).
Артикул КАС Google Scholar
Plate, L. & Marletta, M.A. Зависимая от фосфорилирования дерепрессия с помощью регулятора ответа HnoC в Shewanella oneidensis сигнальная сеть оксида азота. Проц. Натл акад. науч. 110 , E4648–57 (2013).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Риппка, Р., Деруэльес, Дж., Уотербери, Дж. Б., Хердман, М. и Станьер, Р. Ю. Общие присвоения, история штаммов и свойства чистых культур цианобактерий. Микробиология 111 , 1–61 (1979).
Артикул Google Scholar
Henikoff, S. & Henikoff, J.G. Матрицы замещения аминокислот из белковых блоков. Проц. Натл акад. науч. 89 , 10915–10919 (1992).
Артикул ОБЪЯВЛЕНИЯ КАС пабмед ПабМед Центральный Google Scholar
Ссылки на скачивание
Благодарности
Финансовая поддержка этой работы была предоставлена Национальной программой ключевых исследований и разработок Китая (номер гранта 2021YFA0
Финансирование
Финансирование открытого доступа организовано и разрешено Projekt DEAL.
Информация об авторе
Примечания автора
Эти авторы внесли равный вклад: Маркус Циманн, Виктория Рейманн, Яджинг Лян.
Эти авторы совместно руководили этой работой: Tao Zhu, Xuefeng Lu, Wolfgang R. Hess.
Авторы и филиалы
Факультет биологии, Институт биологии III, Генетика и экспериментальная биоинформатика, Фрайбургский университет, Schänzlestr. 1, Фрайбург, D-79104, Германия
Маркус Циманн, Виктория Райманн и Вольфганг Р. Гесс
Институт биоэнергетики и биотехнологий Циндао (QIBEBT), Китайская академия наук, No.189 Songling Road, Циндао, 266101, Китай
Шаньдунский энергетический институт, Циндао, 266101, Китай
Яцзин Лян, Юэ Ши, Хунлей Ма, Тао Чжу и 9 Сюэфэн Лу
Циндаоская лаборатория новой энергии Шаньдун, Циндао, 266101, Китай
Яцзин Лян, Юэ Ши, Хунлей Ма, Тао Чжу и Сюэфэн Лу
Университет Китайской академии наук, Пекин, 100049, Китай
Юман Се, Хуэй Ли, Тао Чжу и Сюэфэн ЛуЛаборатория морской биологии и биотехнологии, Национальная лаборатория морских наук и технологий Циндао, Циндао, 266237, Китай
Сюэфэн Лу
Авторы
- Маркус Циманн
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Viktoria Reimann
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Yajing Liang
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Юэ Ши
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Honglei Ma
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Yuman Xie
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Хуэй Ли
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Tao Zhu
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Xuefeng Lu
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Wolfgang R. Hess
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Contributions
W.R.H., XL, and T.Z. спроектировал исследование. TZ, YX и HL сконструировали мутантные штаммы Anabaena 7120. В.Р. выполнили эксперименты TXTL, анализ микрочипов и нозерн-гибридизацию. М.З. сделал большинство биоинформатических анализов. Ю.С. и Ю.Х. выполнили qRT-PCR и вестерн-блоттинг. Ю.С., Т.З. и Ю.Л. выполнили эксперименты по футпринтингу EMSA и ДНКазы I. Ю.Л. и Х.М. выполнил структурный анализ CvkR. В.Р., М.З., Ю.Л., Т.З. и В.Р.Х. написал статью с участием всех авторов.
Авторы переписки
Переписка с Тао Чжу, Сюэфэн Лу или Вольфганг Р. Гесс.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Рецензирование
Информация о рецензировании
Nature Communications благодарит Ричарда Бреннана, Киру Макарову, Ю Чжан и других анонимных рецензентов за их вклад в рецензирование этой работы. Доступны отчеты рецензентов.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительная информация
Файл рецензирования
Описание дополнительных файлов.0022
Supplementary Data 3
Supplementary Data 4
Supplementary Data 5
Reporting Summary
Source data
Source Data
Rights and permissions
Open Access This article находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете автора(ов) оригинала и источник, предоставите ссылку на лицензии Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатки и разрешения
Об этой статье
Как заморозить свежие овощи
Перейти к рецепту
Сохраняйте вкус лета в течение всего года.
Лето — прекрасная пора для любителей овощей. Сад и фермерский рынок изобилуют свежими продуктами — помидорами, сладкой кукурузой, морковью, болгарским перцем, цуккини до максимума. К сожалению, это не длится. К ноябрю этот поток свежих продуктов иссяк, и они снова вернулись к товарам из супермаркетов.
Но есть и хорошие новости: если у вас есть морозильная камера, вы можете сохранить свежесобранный вкус. Заморозив часть своего урожая сейчас, вы можете наслаждаться домашними овощами всю зиму.
6 шагов по заморозке свежих овощей: давайте начнем с основ с этих 6 шагов
- Подготовьте . Вымойте овощи и, если вы используете паровую корзину, нарежьте их до нужного размера. Я уточню различные требования для разных овощей.
- Бланшировать (варить в кипящей воде) овощи перед замораживанием . Бланширование останавливает ферменты, поддерживающие процесс созревания. Он также помогает избавиться от грязи и бактерий, сохраняет питательные вещества и осветляет цвет. Используйте по крайней мере один галлон воды на фунт овощей. Варите их в воде одну-две минуты. (Примечание: если вы замораживаете помидоры, пропустите этот шаг)
- Сразу после бланширования ненадолго опустите овощи в миску с ледяной водой, пока они полностью не остынут.
- Слейте воду и высушите. Если вы не используете корзину для варки на пару, то на этом этапе отрежьте до желаемого размера.
- Далее вы получите контейнер для морозильной камеры или противень. Сделайте один слой овощей и заморозьте их до твердого состояния.
- После заморозки упакуйте их в пакеты для заморозки или используйте вакуумный упаковщик. Вы не хотите, чтобы они вступали в контакт с воздухом. Это приведет к появлению «неприятных» привкусов к тому времени, когда вы их разморозите.
Полезные советы:
- Заморозьте овощи в течение нескольких часов после их сбора или покупки, чтобы после заморозки они имели наилучший вкус и текстуру. Выбирайте те, которые не повреждены и не имеют надрезов для заморозки.
- Бланширование (кратковременное ошпаривание овощей кипятком или паром) является обязательным для почти всех овощей, подлежащих заморозке. Он останавливает действие ферментов, которые могут привести к потере вкуса, цвета и текстуры. Бланширование очищает поверхность от грязи и организмов, осветляет цвет и помогает замедлить потерю витаминов. Он также делает овощи вялыми или мягче и облегчает их упаковку.
- Время бланширования имеет решающее значение и зависит от овоща и размера . При побледнении стимулирует активность ферментов и хуже, чем при отсутствии побледнения. Чрезмерное бланширование приводит к потере вкуса, цвета, витаминов и минералов. Используйте один галлон воды на фунт подготовленных овощей. Положите овощ в корзину для бланширования и опустите в сильно кипящую воду. Накройте бланшер крышкой. Вода должна снова закипеть в течение 1 минуты, или вы используете слишком много овощей для кипячения воды. Начинайте отсчет времени бланширования, как только вода снова закипит. Поддерживайте высокую температуру в течение времени, указанного в инструкции к овощу, который вы замораживаете.
- Оставьте 1/2 дюйма свободного пространства в верхней части мешков.
Вот разбивка по требованиям различных овощей.
- Сельдерей: Сельдерей входит в список овощей, которые плохо переносят заморозку. Он будет вялым и может не сохранить свой цвет. Это не очень хорошо, если вы ищете хрустящую закуску или хотите добавить немного хруста в салат из макарон. Но если вы ищете аромат сельдерея в жареном блюде, приготовленной запеканке или супе, то это отличный вариант, чтобы добавить к этим блюдам предварительно нарезанный сельдерей. Обрежьте стебли сельдерея, а затем бланшируйте в течение 3 минут и выполните оставшиеся шаги. Если вы хотите мелко нарезать их перед заморозкой, вы можете сделать это или сделать это перед добавлением в блюдо. Они будут прекрасно храниться в вашем холодильнике до года.
- Морковь: Вымойте, очистите и удалите верхушки. Нарежьте тонкими ломтиками, 1/4-дюймовыми кубиками или продольными полосками. Бланшировать небольшие целые моркови в течение 5 минут, нарезать кубиками или ломтиками в течение 2 минут и нарезать вдоль полосками в течение 2 минут. Быстро остудить, процедить, высушить и упаковать, оставив 1/2 дюйма свободного пространства. Запечатать и заморозить. *Убедитесь, что вы используете свежую морковь , иначе она будет иметь резиновый привкус, когда вы будете использовать ее позже. Они останутся свежими в течение года.
- Цуккини/желтая тыква: Вымойте и нарежьте кабачки и/или кабачки до желаемого размера. Тогда вы будете бланшировать всего 2 минуты. Вы не хотите, чтобы он готовил больше, чем это. Перейдите прямо в ледяную баню, а затем заморозьте в один слой на противне. После заморозки вы можете переложить его в пакет для заморозки и снова заморозить. Это похоже на сельдерей в том, что вы не захотите использовать его в качестве гарнира, так как он потеряет часть своей твердости, но его можно добавлять в ваши любимые супы, соусы для спагетти и запеканки. Промаркируйте и поставьте дату на пакете, лучше всего использовать его в течение 3 месяцев, но он продержится дольше в морозильной камере, просто знайте, что чем дольше он там находится, тем он потеряет свой вкус.
- Болгарский перец и лук: Это овощи, которые легче всего заморозить, потому что вам не нужно бланшировать и охлаждать их льдом. Просто нарежьте лук по желанию, а для перца: удалите стебли, семена и мембраны; режь их как хочешь. Затем разложите на подносе, чтобы они не касались друг друга. Поместите их в морозильную камеру, пока они не затвердеют, затем переложите в безопасный для морозильной камеры пакет с застежкой-молнией, выдавив весь воздух, или в вакуумный пакет.
- Кукуруза: Вы можете заморозить целый початок кукурузы, просто отделив его от кожуры, а затем (лично я разрезал бы его пополам, чтобы получить два мини-початка кукурузы), поместив его в пакет для заморозки, выпустив как можно больше воздуха. и замораживание. ИЛИ вы можете бланшировать початки кукурузы в течение 2-3 минут, а затем дать им остыть, пока вы не справитесь с ними. Отрежьте ядра и поместите в литровый пакет, удалите воздух и заморозьте. Они могут храниться до года в морозильной камере.
- Брокколи: Разрежьте брокколи на соцветия и стебли одинакового размера. Затем их нужно хорошо вымыть, чтобы смыть всю грязь. Когда вы бланшируете их, вам нужно будет добавить соль в воду. Бланшируйте в течение 3 минут, а затем переместите на ледяную баню. Затем обсушите их и переложите на противень, застеленный пергаментной бумагой. Заморозить на 1-2 часа, а затем переложить в пакеты для заморозки. Они прослужат до 1 года.
- Картофель : Очистите и вымойте картофель. Затем я разделил их на четыре части и бланшировал 3-5 минут. Поместите их в ледяную баню на 5-10 минут, а затем нарежьте на любой размер, который вы хотите. Упакуйте в пакет для заморозки и удалите как можно больше воздуха.
Я использовал эти удобные держатели для сумок, чтобы сделать этот процесс еще проще!
- Держатели для сумок (партнерская ссылка)
Овощные рецепты, которые мы любим!
- Сэндвичи с круассаном для завтрака, пригодные для заморозки
- (Можно заморозить) Буррито для завтрака
- Кесадилья для завтрака в морозильной камере, 4 вида
- Полезные замороженные банановые лакомства
Ищете другие бесплатные рецепты? Подпишитесь на мою рассылку и следите за всеми последними обновлениями в Facebook, Twitter, Pinterest и Instagram.
Пока нет оценок
Как заморозить свежие овощи
Начнем с 6 шагов, как заморозить свежие овощи. Картофель, кукуруза, морковь, сельдерей, брокколи и многое другое.
Печать Pin This
- celery
- carrots
- bell peppers
- onions
- potatoes
- zucchini
- squash
- corn
- broccoli
Celery : Celery is in the list of veggies that just won не выдерживает заморозки. Он будет вялым и может не сохранить свой цвет. Это не очень хорошо, если вы ищете хрустящую закуску или хотите добавить немного хруста в салат из макарон. Но если вы ищете аромат сельдерея в жареном блюде, приготовленной запеканке или супе, то это отличный вариант, чтобы добавить к этим блюдам предварительно нарезанный сельдерей. Обрежьте стебли сельдерея, а затем бланшируйте в течение 3 минут и выполните оставшиеся шаги.
Если вы хотите мелко нарезать их перед заморозкой, вы можете сделать это или сделать это перед добавлением в блюдо. Они будут прекрасно храниться в вашем холодильнике до года.Морковь : Вымойте, очистите и удалите верхушки. Нарежьте тонкими ломтиками, 1/4-дюймовыми кубиками или продольными полосками. Бланшировать небольшие целые моркови в течение 5 минут, нарезать кубиками или ломтиками 2 минуты и полосками вдоль 2 минуты. Быстро остудить, процедить, высушить и упаковать, оставив 1/2 дюйма свободного пространства. Запечатать и заморозить. * Убедитесь, что вы используете свежую морковь, иначе она будет иметь резиновый привкус, когда вы будете использовать ее позже. Они останутся свежими в течение года.
Цуккини/желтая тыква : Вымойте и нарежьте кабачки и/или кабачки до желаемого размера. Тогда вы будете бланшировать всего 2 минуты. Вы не хотите, чтобы он готовил больше, чем это. Перейдите прямо в ледяную баню, а затем заморозьте в один слой на противне.
После заморозки вы можете переложить его в пакет для заморозки и снова заморозить. Это похоже на сельдерей в том, что вы не захотите использовать его в качестве гарнира, так как он потеряет часть своей твердости, но его можно добавлять в ваши любимые супы, соусы для спагетти и запеканки. Промаркируйте и поставьте дату на пакете, лучше всего использовать его в течение 3 месяцев, но он продержится дольше в морозильной камере, просто знайте, что чем дольше он там находится, тем он потеряет свой вкус.Болгарский перец и лук: Это овощи, которые проще всего заморозить, потому что вам не нужно бланшировать и охлаждать. Просто нарежьте лук по желанию, а для перца: удалите стебли, семена и мембраны; режь их как хочешь. Затем разложите на подносе, чтобы они не касались друг друга. Поместите их в морозильную камеру, пока они не затвердеют, затем переложите в безопасный для морозильной камеры пакет с застежкой-молнией, выдавив весь воздух, или в вакуумный пакет.
Кукуруза: Вы можете заморозить целый початок кукурузы, просто очистив его, а затем (лично я разрежу его пополам, чтобы получить два мини-початка кукурузы), поместив его в пакет для морозильной камеры, выпустив как можно больше воздуха и заморозив. ИЛИ вы можете бланшировать початки кукурузы в течение 2-3 минут, а затем дать им остыть, пока вы не справитесь с ними. Отрежьте ядра и поместите в литровый пакет, удалите воздух и заморозьте. Они могут храниться до года в морозильной камере.
Брокколи: Разрежьте брокколи на соцветия и стебли одинакового размера. Затем их нужно хорошо вымыть, чтобы смыть всю грязь. Когда вы бланшируете их, вам нужно будет добавить соль в воду. Бланшируйте в течение 3 минут, а затем переместите на ледяную баню. Затем обсушите их и переложите на противень, застеленный пергаментной бумагой. Заморозить на 1-2 часа, а затем переложить в пакеты для заморозки. Они прослужат до 1 года.
Картофель: Очистите и вымойте картофель.
Затем я разделил их на четыре части и бланшировал 3-5 минут. Поместите их в ледяную баню на 5-10 минут, а затем нарежьте на любой размер, который вы хотите. Упакуйте в пакет для заморозки и удалите как можно больше воздуха.
Полезные советы:
- Заморозьте овощи в течение нескольких часов после их сбора или покупки, чтобы после заморозки они имели наилучший вкус и текстуру. Выбирайте те, которые не повреждены и не имеют надрезов для заморозки.
- Бланширование (кратковременное ошпаривание овощей кипятком или паром) необходимо практически для всех овощей, подлежащих заморозке. Он останавливает действие ферментов, которые могут привести к потере вкуса, цвета и текстуры. Бланширование очищает поверхность от грязи и организмов, осветляет цвет и помогает замедлить потерю витаминов. Он также делает овощи вялыми или мягче и облегчает их упаковку.
- Время бланширования имеет решающее значение и зависит от овоща и его размера. Недостаточное побледнение стимулирует активность ферментов и хуже, чем отсутствие побледнения. Перебланширование приводит к потере вкуса, цвета, витаминов и минералов. Используйте один галлон воды на фунт подготовленных овощей. Положите овощ в корзину для бланширования и опустите в сильно кипящую воду. Накройте бланшер крышкой. Вода должна снова закипеть в течение 1 минуты, или вы используете слишком много овощей для кипячения воды. Начинайте отсчет времени бланширования, как только вода снова закипит. Поддерживайте высокую температуру в течение времени, указанного в инструкции к овощу, который вы замораживаете.
- Оставьте 1/2 дюйма свободного пространства в верхней части мешков.
Сохраните этот рецепт на потом! Нажмите на сердечко в правом нижнем углу, чтобы сохранить рецепт в свой ящик!
Family Fresh Meals не является диетологом или диетологом, и любая предоставленная информация о питании является только оценочной. Мы рекомендуем прогонять ингредиенты через онлайн-калькулятор питания, если вам нужно проверить какую-либо информацию.
Курс:Гарнир
Кухня:здоровая, вегетарианская
Ключевое слово:советы
Вы приготовили этот рецепт? Отметьте меня в Instagram на @familyfreshmeals или оставьте внизу.
Шаги, которые я использовал, взяты с https://share.upmc.com/2014/06/6-steps-freeze-fresh-vegetables/
Взаимодействие с читателями
Часто задаваемые вопросы о блокировке кредитов | Кража личных данных
Этот информационный бюллетень предназначен для информационных целей и не должен рассматриваться как юридическая консультация или политика штата Нью-Гэмпшир. Если вам нужен совет по конкретному делу, вам следует проконсультироваться с адвокатом или другим экспертом.
- Блокировка кредита
- Часто задаваемые вопросы о заморозке кредита
- Связанные ресурсы
- Бюро защиты прав потребителей и антимонопольного законодательства Дом
Как установить блокировку безопасности?
Могу ли я открыть новые кредитные счета, если мои файлы заморожены?
В чем разница между предупреждением о мошенничестве и блокировкой?
Сколько времени требуется, чтобы заморозка в целях безопасности вступила в силу?
Сколько времени нужно, чтобы снять запрет безопасности?
Что увидит кредитор, запрашивающий мой файл, если он заморожен?
Может ли кредитор получить мой кредитный рейтинг, если мой файл заморожен?
Могу ли я заказать свой собственный кредитный отчет, если мой файл заморожен?
Может ли кто-нибудь увидеть мою кредитную историю, если она заморожена?
Должен ли я заморозить мой файл во всех трех бюро кредитных историй?
Снизит ли заморозка мой кредитный рейтинг?
Может ли работодатель проверить мою биографию, если моя кредитная история заблокирована?
Означает ли замораживание моего файла, что я не буду получать предварительно одобренные кредитные предложения?
Досье моего супруга тоже должно быть заморожено?
Как поставить блокировку безопасности?
Чтобы заморозить кредит, вы должны написать в каждое из трех бюро кредитных историй. Напишите заказным письмом на следующие адреса:
Equifax Security Freeze
Почтовый ящик 105788
Атланта, Джорджия 30348
Experian Security Freeze
а/я 9554
Allen, TX 75013
TransUnion Security Freeze
Почтовый ящик 2000
Chester, PA 19016-2000
И включите следующую информацию:
- Ваше полное имя, включая имя, фамилию, отчество и любой суффикс.
- Любые ранее использовавшиеся следующие имена.
- Ваши текущие полные адреса, включая улицу, любой номер квартиры, город, штат и почтовый индекс.
- Ваши последние полные адреса, включая улицу, любой номер квартиры, город, штат и почтовый индекс.
- Ваш номер социального страхования.
- Дата вашего рождения.
- Если вы стали жертвой кражи личных данных, предоставьте копию полицейского отчета, поданного в местное отделение полиции.
- Если вы не стали жертвой кражи личных данных, приложите платеж в размере 10 долларов США за замораживание.
Могу ли я открыть новые кредитные счета, если мои файлы заморожены?
Да. Если вы хотите открыть новый кредитный счет или получить новый кредит, вы можете отменить блокировку своего кредитного дела. Вы можете поднять его на некоторое время. Или вы можете поднять его для конкретного кредитора. После того, как вы отправите письмо с просьбой о заморозке, каждое кредитное бюро отправит вам личный идентификационный номер (ПИН) в течение 10 рабочих дней с момента получения запроса. Вы также получите инструкции о том, как снять заморозку. Вы можете снять блокировку по телефону, используя свой PIN-код. Бюро кредитных историй должно снять арест в течение трех дней. Плата за временное снятие блокировки составляет 10 долларов США.
В чем разница между предупреждением о мошенничестве и блокировкой?
Предупреждение о мошенничестве — это специальное сообщение в отчете, которое получает кредитный эмитент при проверке кредитного рейтинга потребителя. Он сообщает эмитенту кредита, что со счетом может быть связано мошенничество. Предупреждение о мошенничестве может помочь защитить вас от кражи личных данных. Предупреждение о мошенничестве также может замедлить получение вами нового кредита. Это не должно мешать вам использовать существующие кредитные карты или другие учетные записи. «Заморозка в целях безопасности означает, что потенциальные кредиторы или работодатели не смогут увидеть ваш кредитный файл, проводя проверку биографических данных, если только вы не дадите на это свое согласие. Большинство предприятий не откроют кредитные счета без предварительной проверки кредитной истории потребителя».
Сколько времени требуется, чтобы заморозка в целях безопасности вступила в силу?
Кредитные бюро должны заморозить кредит не позднее чем через пять рабочих дней после получения вашего письменного запроса.
Сколько времени требуется, чтобы запрет безопасности был снят?
Кредитные бюро должны снять блокировку не позднее, чем через три рабочих дня после получения вашего запроса.
Что увидит кредитор, запрашивающий мой файл, если он заморожен?
Кредитор увидит сообщение или код, указывающий, что файл заморожен.
Может ли кредитор получить мой кредитный рейтинг, если мой файл заморожен?
Нет. Кредитор, который запрашивает ваш файл в одном из трех бюро кредитных историй, получит только сообщение или код, указывающий, что файл заморожен.
Могу ли я заказать свой собственный кредитный отчет, если мой файл заморожен?
Да.
Может ли кто-нибудь увидеть мою кредитную историю, если она заморожена?
Если ваш кредитный файл заблокирован в целях безопасности, некоторые лица по-прежнему имеют к нему доступ. Ваш отчет по-прежнему может быть передан вашим существующим кредиторам или коллекторским агентствам, действующим от их имени. Они могут использовать его для просмотра или сбора в вашем аккаунте. Другие кредиторы также могут использовать вашу информацию для предоставления кредита, если только вы не откажетесь от получения таких предложений. Ниже описано, как отказаться от предварительно одобренных кредитных предложений. Правительственные учреждения могут иметь доступ для сбора алиментов или налогов или для расследования мошенничества с Medicaid. Правительственные учреждения также могут иметь доступ в ответ на судебный или административный приказ, повестку в суд или ордер на обыск.
Должен ли я заморозить мой файл во всех трех бюро кредитных историй?
Да. Различные кредитные эмитенты могут использовать разные кредитные бюро. Если вы хотите, чтобы ваш кредитный файл не был просмотрен, вам необходимо заморозить его с помощью Equifax, Experian и TransUnion.
- Образец письма с запросом на блокировку системы безопасности
Снизит ли заморозка мой кредитный рейтинг?
№
Может ли работодатель проверить мою биографию, если моя кредитная история заблокирована?
Нет. Вам придется отменить блокировку, чтобы разрешить проверку биографических данных или подать заявку на страхование, точно так же, как если бы вы подавали заявку на кредит. Вам придется снять замораживание, чтобы разрешить проверку биографических данных, как если бы вы подавали заявку на кредит.
Означает ли замораживание моего файла, что я не буду получать предварительно одобренные кредитные предложения?
Нет. Вы можете отказаться от предварительно одобренных кредитных предложений, позвонив по телефону 888-5OPTOUT (888-567-8688). Или вы можете сделать это онлайн на сайте www.optoutprescreen.com. Это остановит большинство предложений, которые проходят через бюро кредитных историй. Это хорошо на пять лет или вы можете сделать его постоянным.
Досье моего супруга тоже нужно заморозить?
Да. Оба супруга должны заморозить свои отдельные кредитные досье, отправив отдельные письма с просьбой о заморозке, чтобы получить выгоду.
Компьютер зависает при просмотре видео [Исправлено]
Некоторые пользователи сообщают, что их компьютер с Windows зависает или дает сбой при просмотре видео . Неважно, смотрят ли они видео онлайн в своем веб-браузере или в автономном режиме с помощью программного обеспечения медиаплеера, их компьютер либо зависает, либо дает сбой. Если вы столкнулись с той же проблемой на компьютере с Windows 11/10, решения, представленные в этом посте, могут вам помочь.
Если ваш компьютер с Windows 11/10 зависает или зависает при просмотре видео, следующие рекомендации могут помочь вам решить проблему. Прежде чем продолжить, мы рекомендуем вам проверить наличие обновлений Windows. Если доступно обновление, установите его, а затем проверьте, возникает ли проблема.
- Запустите соответствующее средство устранения неполадок
- Откатите драйвер видеокарты или удалите последнюю версию Центра обновления Windows
- Обновите или переустановите драйвер видеокарты
- Устранение неполадок в состоянии чистой загрузки
- Выполнение сканирования SFC и DISM
- Попробуйте другой медиаплеер
- Очистите кэш веб-браузера и файлы cookie
- Попробуйте другой веб-браузер
- Увеличьте виртуальную память
- Создайте новый профиль пользователя Выполните
- 3 90 Обновление на месте
Ниже мы подробно объяснили все эти исправления.
1] Запустите соответствующее средство устранения неполадок
Windows 11/10 имеет встроенные автоматические средства устранения неполадок, которые помогают пользователям устранять проблемы на своих компьютерах. Ваш компьютер дает сбой или зависает каждый раз, когда вы воспроизводите видео. Следовательно, запуск следующего средства устранения неполадок может помочь.
- Средство устранения неполадок воспроизведения видео
- Средство устранения неполадок обслуживания системы
Чтобы запустить средство устранения неполадок воспроизведения видео , выполните указанные ниже действия.
Нажмите на нее, чтобы запустить средство устранения неполадок воспроизведения видео .Действия по запуску средства устранения неполадок обслуживания системы написаны ниже:
- Откройте панель управления.
- Измените режим View by на Large Icons .
- Выберите Поиск и устранение неисправностей .
- Щелкните Система и безопасность .
- Теперь нажмите Обслуживание системы . Это запустит средство устранения неполадок обслуживания системы .
- Когда появится средство устранения неполадок при обслуживании системы, нажмите «Дополнительно» и включите «Автоматически применять ремонт» 9Вариант 3924.
- Если вы видите параметр Запуск от имени администратора , щелкните его, а затем щелкните Далее , чтобы запустить средство устранения неполадок.
Подождите, пока указанные выше средства устранения неполадок завершат процесс устранения неполадок. После этого проверьте, можете ли вы воспроизвести видео или нет. Если нет, попробуйте следующее решение.
Исправление: Проблемы, проблемы и ошибки воспроизведения видео
2] Откатите драйвер видеокарты или удалите последнюю версию Windows Update 9.4296
Вы можете попробовать откатить драйвер видеокарты и посмотреть, поможет ли это. Откройте диспетчер устройств и откройте свойства драйвера видеокарты. Теперь проверьте, доступна ли опция «Откатить». Если да, откатите драйвер. Это действие удалит последнее обновление драйвера.
Если проблема возникла после установки определенного обновления Windows, мы рекомендуем вам удалить последнее обновление Windows и посмотреть, поможет ли оно.
3] Обновите или переустановите драйвер видеокарты
Одной из возможных причин этой проблемы является поврежденный или устаревший драйвер видеокарты. Вам следует обновить драйвер видеокарты. Для этого откройте страницу «Необязательные обновления» в настройках Windows 11/10 и посмотрите, доступно ли там обновление для драйвера вашей видеокарты. Если да, подумайте об установке. Обновление драйвера видеокарты должно решить проблему.
Если это не работает или обновление недоступно, посетите официальный сайт производителя видеокарты и загрузите оттуда последнюю версию драйвера. После этого удалите с компьютера текущую установленную версию драйвера видеокарты с помощью программы удаления драйвера дисплея (DDU). Не волнуйтесь, вы не потеряете свой дисплей после установки драйвера видеокарты. После удаления драйвера запустите установочный файл, загруженный с веб-сайта производителя, чтобы установить драйвер видеокарты вручную.
4] Устранение неполадок в состоянии чистой загрузки
Проблема может быть вызвана конфликтующим фоном или программным обеспечением. Устранив неполадки в вашей системе в состоянии чистой загрузки, вы можете идентифицировать эту программу. В состоянии чистой загрузки Windows запускается только с необходимыми приложениями и службами. Все остальные службы и запускаемые приложения остаются отключенными. После запуска вашей системы в состоянии чистой загрузки воспроизведите видео и посмотрите, не зависает ли оно или не происходит сбой вашей системы. Если нет, вы столкнулись с проблемой из-за проблемного фонового приложения или программного обеспечения.
Чтобы определить проблемное приложение, включите некоторые из отключенных программ в состоянии чистой загрузки, а затем перезагрузите компьютер в обычном режиме. Теперь воспроизведите видео и посмотрите, что произойдет. Если на этот раз ваш компьютер выходит из строя или зависает, виновником является одна из программ, которые вы только что включили. Чтобы определить это, начните отключать программы одну за другой, которые вы только что включили, и воспроизводите видео каждый раз, когда вы отключаете программу. Этот процесс займет время, но поможет вам найти виновника. Как только вы найдете проблемное приложение или программное обеспечение, удалите его и найдите альтернативу.
5] Выполните сканирование SFC и DISM
Одной из возможных причин этой проблемы являются поврежденные файлы образа системы. Существует множество причин повреждения файлов образа системы, таких как скачок напряжения, заражение вредоносным ПО или вирусом и т. д. Если в вашей системе есть какие-либо поврежденные файлы образа системы, сканирование SFC и DISM восстановит их.
После завершения обоих сканирований вы сможете воспроизводить видео без перерыва.
6] Попробуйте другой медиаплеер
В Интернете доступно несколько бесплатных программ для медиаплееров. Вы можете загрузить и установить эти бесплатные медиаплееры в своей системе для воспроизведения видео.
7] Очистите кэш веб-браузера и файлы cookie.
Если воспроизведение видео в веб-браузере приводит к зависанию или сбою компьютера, возможно, поврежден кэш или данные файлов cookie. Очистите кеш и данные cookie вашего веб-браузера и посмотрите, поможет ли это. Для этого нажмите клавиши Ctrl + Shift + Delete. Этот ярлык откроет Окно Clear Browsing Data в большинстве веб-браузеров. Теперь выберите файлы cookie и кеш и удалите их.
8] Попробуйте другой веб-браузер
Воспроизведите видео в другом веб-браузере. Это позволит вам узнать, возникает ли проблема в конкретном веб-браузере или нет. Если воспроизведение видео не зависает или не приводит к сбою компьютера, перезагрузите браузер, с которым вы столкнулись. Процесс сброса некоторых популярных веб-браузеров описан ниже:
- Чтобы сбросить настройки Chrome по умолчанию, откройте настройки Chrome, затем откройте Расширенные настройки . Прокрутите вниз, и вы увидите опцию « Восстановить настройки до исходных значений по умолчанию ».
- Чтобы сбросить или обновить Firefox, откройте Настройки Firefox, затем перейдите в « Справка > Информация об устранении неполадок ». Теперь нажмите кнопку Обновить Firefox .
- Чтобы восстановить настройки Microsoft Edge по умолчанию, откройте Edge и перейдите в «Настройки > Сбросить настройки ».
9] Увеличение виртуальной памяти
Другим эффективным решением этой проблемы является увеличение виртуальной памяти. Увеличение виртуальной памяти может повысить производительность системы. Если это решит проблему, обновите физическую оперативную память. Если этот метод не устраняет проблему, отмените внесенные изменения и попробуйте следующее решение.
10] Создайте новую учетную запись пользователя
Иногда проблема возникает с определенной учетной записью пользователя. Вы можете проверить это, создав новую учетную запись пользователя. Откройте настройки Windows 11/10 и создайте новую учетную запись локального администратора. После создания новой учетной записи пользователя выйдите из своей текущей учетной записи и войдите в новую учетную запись пользователя, которую вы только что создали. Теперь воспроизведите видео и посмотрите, что произойдет.
11] Выполните обновление на месте
Если ни одно из приведенных выше решений не помогло вам, выполните обновление на месте. Обновление на месте устанавливает операционную систему Windows без удаления существующей. Он также восстанавливает систему пользователя. Хотя обновление на месте не удаляет данные с вашего компьютера, мы рекомендуем сделать резервную копию всех ваших данных на внешнем жестком диске, прежде чем продолжить.
Чтение : Windows аварийно завершает работу или зависает даже в безопасном режиме.
Почему мои видео постоянно зависают?
Может быть много причин, по которым ваше видео зависает на компьютере с Windows. Если медиаплеер, который вы используете для воспроизведения видео, не поддерживает кодеки, он может зависнуть. Другой причиной этой проблемы являются поврежденные файлы образа системы. В таком случае вам следует запустить сканирование SFC и DISM, чтобы решить проблему.
Если у вас возникли проблемы с зависанием при воспроизведении видео в веб-браузере, удалите его файлы cookie и кэш-данные. Кроме того, проверьте, возникает ли проблема в других веб-браузерах. Если нет, вам необходимо перезагрузить браузер, с которым у вас возникла проблема.
Иногда проблемное фоновое приложение вызывает проблемы на устройстве Windows. Вы можете определить эту программу, устранив неполадки на компьютере в состоянии чистой загрузки.
Вам также следует обновить драйвер видеокарты и посмотреть, поможет ли это. Если проблема возникает на определенном медиаплеере, переключение на другой медиаплеер решит проблему.
Почему ПК внезапно зависает?
Если ваш ПК с Windows внезапно зависает, файлы образа системы могут быть повреждены. В этом случае поможет запуск сканирования SFC и DISM.
Если проблема возникает часто, это может указывать на неисправность ОЗУ. Вы можете проверить состояние вашей оперативной памяти с помощью средства диагностики памяти. Если ваша оперативная память исправна и ваш компьютер зависает при запуске нескольких программ, вам следует обновить физическую оперативную память.
Надеюсь, это поможет.
Читать дальше : Компьютер Windows зависает при установке графического драйвера.
заморозка — Liquipedia Counter-Strike Wiki
Следите за нами в Твиттере @LiquipediaCS, если хотите быть в курсе всех событий Counter-Strike!
Из Liquipedia Counter-Strike Wiki
Давид « замерзший » Чернанский (родился 18 июля 2002 г.) — словацкий профессиональный игрок Counter-Strike: Global Offensive для MOUZ.
Содержимое
- 1 Механизм и настройки
- 2 достижения
- 3 Общая информация
- 4 Особенности
- 4.1 HLTV
- 4.2 Другое
- 5 Интервью
- 6 Галерея
- 7 Ссылки
Шестерня и настройки
Advanced
DeathAdder V2 Pro
9| Mouse Settings [1] (список) (вычислить) Простой | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Мышь | КРИВАЦИЯ | Поход | Установка мыши | 9443.4540 | Масштаб | | ||||||
| Razer DeathAdder V2 Pro | 1003 MPI 0.658 deg/mm | 21.5 in/rev 54.7 cm/rev | 400 DPI @ 1000 Гц | 1,9 | 0,8 | On | ||||||
| Обновлено от 23 января 2021 г.)1397 | ||||||||||||
| Оборудование [1] |
|---|
| Монитор | Частота обновления | Разрешение в игре | Масштабирование | |
|---|---|---|---|---|
| ZOWIE XL2546 | 240 Гц | 1280×960 | Растянутый | |
Примечание: Покупки, сделанные по приведенным выше розничным ссылкам, могут принести Liquipedia небольшую комиссию.
| Crosshair Settings [2] (list of) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Style | Size | Thickness | Sniper | Gap | Outline | Dot | Color | Alpha | |||
4 | 3 | 0 | 1 | -3 | No | No ( 0 ) | Green ( 1 ) | 255 | |||
| Updated as of 2022- 04-17 (321 день назад) | |||||||||||
| Нажмите «Развернуть», чтобы скопировать и вставить общий код и консольные команды. | |
|---|---|
| Sharecode | CSGO-jmBsb-LKp6T-aqzyY-f6Uaj-aGUSH |
| Команды | cl_crosshairalpha 255; cl_crosshaircolor 1; cl_crosshairdot 0; cl_crosshairgap -3; cl_crosshairsize 3; cl_crosshairstyle 4; cl_crosshairusealpha 1; cl_crosshairthickness 0; cl_crosshair_sniper_width 1; cl_crosshair_outlinethickness 1; cl_crosshair_drawoutline 0; |
Достижения4869 Tournament
0005- назван Magisk и Guardian в качестве потенциального игрока 2017 года. [3] [4]
- Занял 17-е место среди лучших игроков 2022 года по версии HLTV. [5]
- Был назван самым ценным игроком третьего сезона «Точки воспламенения» по версии HLTV. [6]
Особенности[править]
HLTV[править]
- 28 марта 2017 | Frozen vs. Singularity от brcho HLTV на ESEA Season 24: Open Division — Europe
- 12.11.2017 | Frozen vs. PRIDE от brcho HLTV на Europe Minor Championship — Boston 2018
- 23 декабря 2019 | заморожено по сравнению с Vitality от FILQ из HLTV на EPICENTER 2019
- 04.05.2020 | заморожено по сравнению с G2 от MAKKU из HLTV на ESL One: Road to Rio — Europe
- 19.09.2020 | Frozen vs. Vitality от MAKKU из HLTV на ESL Pro League Season 12: Europe
- 29.10.2021 | Fзамороженный получает туз против BIG от FILQ HLTV на PGL Major Stockholm 2021
Другое[править]
- 12.11.2015 | Frozen: Нехраю на фраги, храю про тим [Frozen: Я не играю за фраги, я играю за команду] by недостаточный из PLAYzone на Mistrovství ČR 2015
- 13.07.2016 | заморожено для RoG FINALS 10 [заморожено после RoG FINALS 10] от Hitpoint.cz на ASUS Finals 10
- 2017-03-02 | Рожовор с Фрозеном! [Интервью с Frozen!] Дамиан Есенски из E-GAMES.SK
- 15.05.2017 | Лучший профессиональный консультант по статистике Frozen (14): Vreckové od rodičov už nepotrebujem [Самый молодой профессиональный игрок в компьютерные игры Frozen (14): Мне больше не нужны карманные деньги от родителей] Марсель Марцишяк из TVnoviny.sk
- 05.01.2018 | заморожен из-за возрастных ограничений: «Я ничего не могу с этим поделать, мне просто нужно подождать» от tolkienfanatic из HLTV на ESEA Season 26: Global Challenge
- 10 августа 2018 | Рожовор с Frozen [Интервью с Frozen] от NVIDIA
- 11. 11.2018 | заморожено: Титул для меня znamená víc než MVP [заморожено: титул значит для меня больше, чем MVP] от MrKenny из PLAYzone на Mistrovství ČR 2018
- 2019-01-31 | TOP 10 CZSK: заморожено (#4) от huhy из PLAYzone
- 04.05.2019 | заморожен: «В основном, все, что я делаю, это сплю, хожу в школу и играю в CS» от профессора HLTV на IEM Sydney 2019
- 07.07.2019 | Frozen (mousesports) : «J’aime juste le jeu» [frozen (mousesports) : «Мне просто нравится игра»] VaKaRm на ESL One: Cologne 2019
- 20.07.2019 | заморожен: «Я всегда мечтал сыграть на мейджоре и иметь свои собственные стикеры»
- 30.08.2019 | Дэвид «Frozen» Чернанский о Starladder CS:GO Major Berlin [Дэвид «Frozen» Чернанский о Starladder CS:GO Major Berlin] by Chance на ESL One: Cologne 2019
- 21 сентября 2019 | «Rengeteg jó játékos van az országban, de valami nem klappol» — заморожено [«В Венгрии так много хороших игроков, хотя что-то не подходит» — заморожено] Бен из Esport 1 на V4 Future Sports Festival — Будапешт 2019
- 2019-10-02 | заморожен из-за проигрыша NiP: «Я сказал ребятам, что карта у меня с собой» ТОП-10 ЧСК: заморожено (#1) от huhy из PLAYzone
- 2021-02-02 | Frozen: Celý rok bol viac-menej стереотип – есть, спать, CS:GO
- 08.
05.2021 | «Думаю, мы будем опасны» — заморожено от Фрейи из Flashpoint в 3 сезоне Flashpoint - 08.
- 05.02.2022 | Som veľmi motivovaný a úprimne zvedavý, hlásí frosted premierou letošní premiérou [Я очень мотивирован и искренне любопытен, заморожен говорит перед премьерой в этом году] от huhy из PLAYzone
- 06.07.2022 | замерз: Думаю, мы сможем удивить на групповом этапе от Pley на IEM Cologne 2022
- 08.07.2022 | заморожены после того, как звенят героически: «Det känns fucking fantastiskt» [застыл после победы над Heroic: «Это чертовски потрясающе»] от Hallonkraem из Fragbite на IEM Cologne 2022
- 14.07.2022 | «Если бы я не сделал это в CS, я бы, наверное, попробовал это в LoL.» | 20 минут с замороженным от МОУЗ
Галерея «замороженный». параметры
